PHORATE First Draft Prepared by P.K. Gupta Toxicology Consulting
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
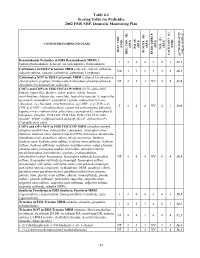
2002 NRP Section 6, Tables 6.1 Through
Table 6.1 Scoring Table for Pesticides 2002 FSIS NRP, Domestic Monitoring Plan } +1 0.05] COMPOUND/COMPOUND CLASS * ) (EPA) (EPA) (EPA) (EPA) (EPA) (FSIS) (FSIS) PSI (P) TOX.(T) L-1 HIST. VIOL. BIOCON. (B) {[( (2*R+P+B)/4]*T} REG. CON. (R) * ENDO. DISRUP. LACK INFO. (L) LACK INFO. {[ Benzimidazole Pesticides in FSIS Benzimidazole MRM (5- 131434312.1 hydroxythiabendazole, benomyl (as carbendazim), thiabendazole) Carbamates in FSIS Carbamate MRM (aldicarb, aldicarb sulfoxide, NA44234416.1 aldicarb sulfone, carbaryl, carbofuran, carbofuran 3-hydroxy) Carbamates NOT in FSIS Carbamate MRM (carbaryl 5,6-dihydroxy, chlorpropham, propham, thiobencarb, 4-chlorobenzylmethylsulfone,4- NT 4 1 3 NV 4 4 13.8 chlorobenzylmethylsulfone sulfoxide) CHC's and COP's in FSIS CHC/COP MRM (HCB, alpha-BHC, lindane, heptachlor, dieldrin, aldrin, endrin, ronnel, linuron, oxychlordane, chlorpyrifos, nonachlor, heptachlor epoxide A, heptachlor epoxide B, endosulfan I, endosulfan I sulfate, endosulfan II, trans- chlordane, cis-chlordane, chlorfenvinphos, p,p'-DDE, p, p'-TDE, o,p'- 3444NV4116.0 DDT, p,p'-DDT, carbophenothion, captan, tetrachlorvinphos [stirofos], kepone, mirex, methoxychlor, phosalone, coumaphos-O, coumaphos-S, toxaphene, famphur, PCB 1242, PCB 1248, PCB 1254, PCB 1260, dicofol*, PBBs*, polybrominated diphenyl ethers*, deltamethrin*) (*identification only) COP's and OP's NOT in FSIS CHC/COP MRM (azinphos-methyl, azinphos-methyl oxon, chlorpyrifos, coumaphos, coumaphos oxon, diazinon, diazinon oxon, diazinon met G-27550, dichlorvos, dimethoate, dimethoate -

Chemical Name Federal P Code CAS Registry Number Acutely
Acutely / Extremely Hazardous Waste List Federal P CAS Registry Acutely / Extremely Chemical Name Code Number Hazardous 4,7-Methano-1H-indene, 1,4,5,6,7,8,8-heptachloro-3a,4,7,7a-tetrahydro- P059 76-44-8 Acutely Hazardous 6,9-Methano-2,4,3-benzodioxathiepin, 6,7,8,9,10,10- hexachloro-1,5,5a,6,9,9a-hexahydro-, 3-oxide P050 115-29-7 Acutely Hazardous Methanimidamide, N,N-dimethyl-N'-[2-methyl-4-[[(methylamino)carbonyl]oxy]phenyl]- P197 17702-57-7 Acutely Hazardous 1-(o-Chlorophenyl)thiourea P026 5344-82-1 Acutely Hazardous 1-(o-Chlorophenyl)thiourea 5344-82-1 Extremely Hazardous 1,1,1-Trichloro-2, -bis(p-methoxyphenyl)ethane Extremely Hazardous 1,1a,2,2,3,3a,4,5,5,5a,5b,6-Dodecachlorooctahydro-1,3,4-metheno-1H-cyclobuta (cd) pentalene, Dechlorane Extremely Hazardous 1,1a,3,3a,4,5,5,5a,5b,6-Decachloro--octahydro-1,2,4-metheno-2H-cyclobuta (cd) pentalen-2- one, chlorecone Extremely Hazardous 1,1-Dimethylhydrazine 57-14-7 Extremely Hazardous 1,2,3,4,10,10-Hexachloro-6,7-epoxy-1,4,4,4a,5,6,7,8,8a-octahydro-1,4-endo-endo-5,8- dimethanonaph-thalene Extremely Hazardous 1,2,3-Propanetriol, trinitrate P081 55-63-0 Acutely Hazardous 1,2,3-Propanetriol, trinitrate 55-63-0 Extremely Hazardous 1,2,4,5,6,7,8,8-Octachloro-4,7-methano-3a,4,7,7a-tetra- hydro- indane Extremely Hazardous 1,2-Benzenediol, 4-[1-hydroxy-2-(methylamino)ethyl]- 51-43-4 Extremely Hazardous 1,2-Benzenediol, 4-[1-hydroxy-2-(methylamino)ethyl]-, P042 51-43-4 Acutely Hazardous 1,2-Dibromo-3-chloropropane 96-12-8 Extremely Hazardous 1,2-Propylenimine P067 75-55-8 Acutely Hazardous 1,2-Propylenimine 75-55-8 Extremely Hazardous 1,3,4,5,6,7,8,8-Octachloro-1,3,3a,4,7,7a-hexahydro-4,7-methanoisobenzofuran Extremely Hazardous 1,3-Dithiolane-2-carboxaldehyde, 2,4-dimethyl-, O- [(methylamino)-carbonyl]oxime 26419-73-8 Extremely Hazardous 1,3-Dithiolane-2-carboxaldehyde, 2,4-dimethyl-, O- [(methylamino)-carbonyl]oxime. -

Lifetime Organophosphorous Insecticide Use Among Private Pesticide Applicators in the Agricultural Health Study
Journal of Exposure Science and Environmental Epidemiology (2012) 22, 584 -- 592 & 2012 Nature America, Inc. All rights reserved 1559-0631/12 www.nature.com/jes ORIGINAL ARTICLE Lifetime organophosphorous insecticide use among private pesticide applicators in the Agricultural Health Study Jane A. Hoppin1, Stuart Long2, David M. Umbach3, Jay H. Lubin4, Sarah E. Starks5, Fred Gerr5, Kent Thomas6, Cynthia J. Hines7, Scott Weichenthal8, Freya Kamel1, Stella Koutros9, Michael Alavanja9, Laura E. Beane Freeman9 and Dale P. Sandler1 Organophosphorous insecticides (OPs) are the most commonly used insecticides in US agriculture, but little information is available regarding specific OP use by individual farmers. We describe OP use for licensed private pesticide applicators from Iowa and North Carolina in the Agricultural Health Study (AHS) using lifetime pesticide use data from 701 randomly selected male participants collected at three time periods. Of 27 OPs studied, 20 were used by 41%. Overall, 95% had ever applied at least one OP. The median number of different OPs used was 4 (maximum ¼ 13). Malathion was the most commonly used OP (74%) followed by chlorpyrifos (54%). OP use declined over time. At the first interview (1993--1997), 68% of participants had applied OPs in the past year; by the last interview (2005--2007), only 42% had. Similarly, median annual application days of OPs declined from 13.5 to 6 days. Although OP use was common, the specific OPs used varied by state, time period, and individual. Much of the variability in OP use was associated with the choice of OP, rather than the frequency or duration of application. -

Quantum Chemical Study of the Thermochemical Properties of Organophosphorous Compounds A
QUANTUM CHEMICAL STUDY OF THE THERMOCHEMICAL PROPERTIES OF ORGANOPHOSPHOROUS COMPOUNDS A. Khalfa, M. Ferrari, R. Fournet, B. Sirjean, L. Verdier, Pierre-Alexandre Glaude To cite this version: A. Khalfa, M. Ferrari, R. Fournet, B. Sirjean, L. Verdier, et al.. QUANTUM CHEMICAL STUDY OF THE THERMOCHEMICAL PROPERTIES OF ORGANOPHOSPHOROUS COMPOUNDS. Journal of Physical Chemistry A, American Chemical Society, 2015, 119 (42), pp.10527-10539. 10.1021/acs.jpca.5b07071. hal-01241498 HAL Id: hal-01241498 https://hal.archives-ouvertes.fr/hal-01241498 Submitted on 10 Dec 2015 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. QUANTUM CHEMICAL STUDY OF THE THERMOCHEMICAL PROPERTIES OF ORGANOPHOSPHOROUS COMPOUNDS A. Khalfa, M. Ferrari1, R. Fournet1, B. Sirjean1, L. Verdier2, P.A. Glaude1 1Laboratoire Réactions et Génie des Procédés, Université de Lorraine, CNRS, 1 rue Grandville, BP 20451, 54001 NANCY Cedex, France, 2DGA Maîtrise NRBC, Site du Bouchet, 5 rue Lavoisier, BP n°3, 91710 Vert le Petit, France Abstract Organophosphorous compounds are involved in many toxic compounds such as fungicides, pesticides, or chemical warfare nerve agents. The understanding of the decomposition chemistry of these compounds in the environment is largely limited by the scarcity of thermochemical data. -

Fate and Effects of Azinphos-Methyl in a Flow-Through Wetland in South
Environ. Sci. Technol. 2003, 37, 2139-2144 preventing it from entering downstream aquatic habitats (1, Fate and Effects of Azinphos-Methyl 2). The implementation of retention ponds in agricultural in a Flow-Through Wetland in South watersheds was mentioned by Scott et al. (3) as one strategy to reduce the amount and toxicity of runoff-related insecticide Africa pollution discharging into estuaries. The usefulness of aquatic plants for removal of insecticides from water has been shown ,² ² in an indoor microcosm study (4), and the effects of the RALF SCHULZ,* CHRISTINA HAHN, organophosphate phorate have been assessed using littoral ERIN R. BENNETT,² mesocosms in South Dakota wetlands (5). However, infor- JAMES M. DABROWSKI,² mation about the fate or effects of spray drift-borne GERALDINE THIERE,² AND SUE K. C. PEALL³ insecticide input in constructed wetlands is limited. Processes important for removal of nonpoint-source Department of Zoology, Private Bag X1, University of pesticide pollution in wetlands may include adsorption, Stellenbosch, Matieland 7602, South Africa, and Forensic Chemistry Laboratory, Department of Health, decomposition, hydrolysis, microbial metabolism, photolysis, Cape Town 8000, South Africa and volatilization (6). The macrophytes present in the wetland may play an important role in providing an increased surface area for sorption as well as for microbial activity (7). Further- more, they may contribute directly to metabolism (8). Our knowledge about the effectiveness of constructed Spray drift is an important route for nonpoint-source wetlands in retaining agricultural nonpoint-source pesticide pesticide pollution of aquatic habitats (9, 10). Specifically, pollution is limited. A 0.44-ha vegetated wetland built orchard applications result in a large amount of drift due to small droplet size and the trajectory of release (11). -

Code Chemical P026 1-(O-Chlorophenyl)Thiourea P081 1
Code Chemical P026 1-(o-Chlorophenyl)thiourea P081 1,2,3-Propanetriol, trinitrate (R) P042 1,2-Benzenediol, 4-[1-hydroxy-2-(methylamino)ethyl]-, (R)- P067 1,2-Propylenimine P185 1,3-Dithiolane-2-carboxaldehyde, 2,4-dimethyl-, O- [(methylamino)- carbonyl]oxime 1,4,5,8-Dimethanonaphthalene, 1,2,3,4,10,10-hexa- chloro-1,4,4a,5,8,8a,-hexahydro-, P004 (1alpha,4alpha, 4abeta,5alpha,8alpha,8abeta)- 1,4,5,8-Dimethanonaphthalene, 1,2,3,4,10,10-hexa- chloro-1,4,4a,5,8,8a-hexahydro-, P060 (1alpha,4alpha, 4abeta,5beta,8beta,8abeta)- P002 1-Acetyl-2-thiourea P048 2,4-Dinitrophenol P051 2,7:3,6-Dimethanonaphth [2,3-b]oxirene, 3,4,5,6,9,9 -hexachloro-1a,2,2a,3,6,6a,7,7a- octahydro-, (1aalpha,2beta,2abeta,3alpha,6alpha,6abeta,7 beta, 7aalpha)-, & metabolites 2,7:3,6-Dimethanonaphth[2,3-b]oxirene, 3,4,5,6,9,9- hexachloro-1a,2,2a,3,6,6a,7,7a- P037 octahydro-, (1aalpha,2beta,2aalpha,3beta,6beta,6aalpha,7 beta, 7aalpha)- P045 2-Butanone, 3,3-dimethyl-1-(methylthio)-, O-[methylamino)carbonyl] oxime P034 2-Cyclohexyl-4,6-dinitrophenol 2H-1-Benzopyran-2-one, 4-hydroxy-3-(3-oxo-1- phenylbutyl)-, & salts, when present at P001 concentrations greater than 0.3% P069 2-Methyllactonitrile P017 2-Propanone, 1-bromo- P005 2-Propen-1-ol P003 2-Propenal P102 2-Propyn-1-ol P007 3(2H)-Isoxazolone, 5-(aminomethyl)- P027 3-Chloropropionitrile P047 4,6-Dinitro-o-cresol, & salts P059 4,7-Methano-1H-indene, 1,4,5,6,7,8,8-heptachloro- 3a,4,7,7a-tetrahydro- P008 4-Aminopyridine P008 4-Pyridinamine P007 5-(Aminomethyl)-3-isoxazolol 6,9-Methano-2,4,3-benzodioxathiepin, 6,7,8,9,10,10- -

Parathion-Methyl
FAO SPECIFICATIONS AND EVALUATIONS FOR PLANT PROTECTION PRODUCTS PARATHION-METHYL O,O-dimethyl O-4-nitrophenyl phosphorothioate 2001 TABLE OF CONTENTS PARATHION-METHYL Page DISCLAIMER 3 INTRODUCTION 4 PART ONE 5 SPECIFICATIONS FOR PARATHION-METHYL PARATHION-METHYL INFORMATIONERROR! BOOKMARK NOT DEFINED. PARATHION-METHYL TECHNICAL MATERIAL 6 PARATHION-METHYL TECHNICAL CONCENTRATE 8 PARATHION-METHYL EMULSIFIABLE CONCENTRATE 10 PART TWO 13 2001 EVALUATION REPORT ON PARATHION-METHYL 14 Page 2 of 31 PARATHION-METHYL SPECIFICATIONS 2001 Disclaimer1 FAO specifications are developed with the basic objective of ensuring that pesticides complying with them are satisfactory for the purpose for which they are intended so that they may serve as an international point of reference. The specifications do not constitute an endorsement or warranty of the use of a particular pesticide for a particular purpose. Neither do they constitute a warranty that pesticides complying with these specifications are suitable for the control of any given pest, or for use in a particular area. Owing to the complexity of the problems involved, the suitability of pesticides for a particular application must be decided at the national or provincial level. Furthermore, the preparation and use of pesticides complying with these specifications are not exempted from any safety regulation or other legal or administrative provision applicable thereto. FAO shall not be liable for any injury, loss, damage or prejudice of any kind that may be suffered as a result of the preparation, transportation, sale or use of pesticides complying with these specifications. Additionally, FAO wishes to alert users of specifications to the fact that improper field mixing and/or application of pesticides can result in either a lowering or complete loss of efficacy. -

Pesticides and Toxic Substances
UNITED STATES ENVIRONMENTAL PROTECTION AGENCY WASHINGTON D.C., 20460 OFFICE OF PREVENTION, PESTICIDES AND TOXIC SUBSTANCES MEMORANDUM DATE: July 31, 2006 SUBJECT: Finalization of Interim Reregistration Eligibility Decisions (IREDs) and Interim Tolerance Reassessment and Risk Management Decisions (TREDs) for the Organophosphate Pesticides, and Completion of the Tolerance Reassessment and Reregistration Eligibility Process for the Organophosphate Pesticides FROM: Debra Edwards, Director Special Review and Reregistration Division Office of Pesticide Programs TO: Jim Jones, Director Office of Pesticide Programs As you know, EPA has completed its assessment of the cumulative risks from the organophosphate (OP) class of pesticides as required by the Food Quality Protection Act of 1996. In addition, the individual OPs have also been subject to review through the individual- chemical review process. The Agency’s review of individual OPs has resulted in the issuance of Interim Reregistration Eligibility Decisions (IREDs) for 22 OPs, interim Tolerance Reassessment and Risk Management Decisions (TREDs) for 8 OPs, and a Reregistration Eligibility Decision (RED) for one OP, malathion.1 These 31 OPs are listed in Appendix A. EPA has concluded, after completing its assessment of the cumulative risks associated with exposures to all of the OPs, that: (1) the pesticides covered by the IREDs that were pending the results of the OP cumulative assessment (listed in Attachment A) are indeed eligible for reregistration; and 1 Malathion is included in the OP cumulative assessment. However, the Agency has issued a RED for malathion, rather than an IRED, because the decision was signed on the same day as the completion of the OP cumulative assessment. -

Phorate Interim AEGL Document
1 2 3 4 ACUTE EXPOSURE GUIDELINE LEVELS (AEGLs) 5 FOR 6 PHORATE 7 (CAS Reg. No. 298-02-2) 8 9 INTERIM 10 11 12 13 14 15 16 17 18 PHORATE Interim 09-2009; Page 2 of 30 1 2 ACUTE EXPOSURE GUIDELINE LEVELS (AEGLs) 3 FOR 4 PHORATE 5 (CAS Reg. No. 298-02-2) 6 7 8 9 10 INTERIM 11 12 13 14 15 16 17 18 19 20 21 22 23 PHORATE Interim 09-2009; Page 3 of 30 1 2 3 PREFACE 4 5 Under the authority of the Federal Advisory Committee Act (FACA) P. L. 92-463 of 6 1972, the National Advisory Committee for Acute Exposure Guideline Levels for Hazardous 7 Substances (NAC/AEGL Committee) has been established to identify, review and interpret 8 relevant toxicologic and other scientific data and develop AEGLs for high priority, acutely toxic 9 chemicals. 10 11 AEGLs represent threshold exposure limits for the general public and are applicable to 12 emergency exposure periods ranging from 10 minutes to 8 hours. Three levels C AEGL-1, 13 AEGL-2 and AEGL-3 C are developed for each of five exposure periods (10 and 30 minutes, 1 14 hour, 4 hours, and 8 hours) and are distinguished by varying degrees of severity of toxic effects. 15 The three AEGLs are defined as follows: 16 17 AEGL-1 is the airborne concentration (expressed as parts per million or milligrams per 18 cubic meter [ppm or mg/m3]) of a substance above which it is predicted that the general 19 population, including susceptible individuals, could experience notable discomfort, irritation, or 20 certain asymptomatic, non-sensory effects. -

Visualisation of DCP, a Nerve Agent Mimic, in Catfish Brain by a Simple
www.nature.com/scientificreports OPEN Visualisation of DCP, a nerve agent mimic, in Catfsh brain by a simple chemosensor Received: 25 October 2017 Himadri Sekhar Sarkar 1, Ayndrila Ghosh1, Sujoy Das1, Pulak Kumar Maiti2, Sudipta Maitra3, Accepted: 9 February 2018 Sukhendu Mandal 2 & Prithidipa Sahoo 1 Published: xx xx xxxx A chemosensor, 3-aminophenol-based rhodamine conjugate (ARC) has been developed for visualisation of diethylchlorophosphate (DCP), mimic of a chemical warfare agent, in Catfsh brain. The simple detection of DCP by “turn-on” fuorescence property of the chemosensor makes it unique for easy and rapid in vivo and in vitro detection of DCP with the detection limit of 5.6 nM. 1995, Te terrorist attack on Tokyo subway introduces the whole world with a new threat to mankind- Chemical Warfare Agents (CWAs)1–3. Te simple organophosphates, present in pesticides eventually become more pop- ular chemical weapon due to its very simple method of manufacturing, availability, and low cost along with its dispensability. Despite of its common use as a pesticide, various analogues of such organophosphates are found to be very potent nerve agent, which irreversibly damage functions of nerve cells. Among all the highly toxic, volatile nerve agents, Sarin (GB), Soman (GD) and Tabun (GA) are of most common. If the nerve agents are being inhaled or absorbed through skin, their reactive phosphate group irreversibly react with the hydroxyl group of cellular acetylcholinesterase, which is responsible for breaking down the acetylcholine neurotransmitter, leads to its inactivation. Te fatal consequences are neurological imbalance at cholinergic synapses, failure of several organs, paralysis of central nervous system and rapid death4,5. -

Interactions of Acephate, Glyphosate, Monocrotophos and Phorate with Bovine Serum Albumin
Short Communications Interactions of Acephate, Glyphosate, Monocrotophos and Phorate with Bovine Serum Albumin V. KUMAR* AND S. SINGH1 Regional Ayurveda Research Institute for Drug Development, Amkhoh, Gwalior-474 009, 1Department of Biotechnology, Lovely Professional University, Punjab-144 111, India Kumar, et al.: Interactions of Organophosphate Pesticides with BSA Binding of pesticides to serum albumin significantly influence their absorption, metabolism, distribution and excretion. In the present study, interactions of acephate, glyphosate, monocrotophos and phorate with bovine serum albumin were explored employing the UV/Vis and Fourier-transform infrared spectroscopy methods. The observed values of binding constant for titled pesticides were phorate (1.85×107 M-1)>acephate (3.93×104 M-1)>glyphosate (1.31×104 M-1)>monocrotophos (1.12×104 M-1). Results have shown that out of four pesticides, phorate could show strong interactions with bovine serum albumin. Key words: Pesticides, UV/Vis and FTIR, BSA, binding constant Crop protection has become necessary in order to Most importantly, our special interest in the pesticides increase the food production and use of pesticides of different classes allows us to describe the interaction has become unavoidable act for that purpose[1-5]. of acephate, glyphosate, monocrotophos and phorate Pesticides are considered as an integral part of modern with bovine serum albumin (BSA). agriculture[6-10]. At the same time, food safety issues All reagents and solvents used were of the commercial have gained considerable attention due to its effects quality and were used without purification. Technical on environment and human health. Pesticides used to grade level acephate, glyphosate, monocrotophos and protect the crops or foods at various stages like sowing phorate were obtained as gift samples from Gautami to storage, and these synthetic chemicals are allow Ltd., Andhra Pradesh, India and BSA was purchased to enter into human body through different modes of from Loba Chemie Pvt. -

Organophosphate and N-Methyl Carbamate 03-12-2010.Xlsx
Product Name EPA Reg. No. Ingredients Registrant Signal Word AgriSolutions Actellic 5E 1381‐170 Pirimiphos‐methyl Winfield Solutions, LLC Danger Insecticide AgriSolutions Dimate 4E 9779‐273 Dimethoate Winfield Solutions, LLC Warning AgriSolutions Dimate 4EC 51036‐110‐9779 Dimethoate Winfield Solutions, LLC Warning Systemic Insecticide AgriSolutions Malathion 5 9779‐5 Malathion Winfield Solutions, LLC Warning AgriSolutions Phorate 20G 9779‐293 Phorate Winfield Solutions, LLC Danger/Poison AgriSolutions Yuma 4E Insecticide 62719‐220‐1381 Chlorpyrifos Winfield Solutions, LLC Warning Azinphosmethyl 50W Soluble 66330‐233 AZM (Azinphos‐methyl) Arysta LifeScience North America, LLC Danger/Poison Carzol SP Insecticide in Water 10163‐265 Formetanate hydrochloride Gowan Company Danger/Poison Soluble Packaging Chemathoate Technical 4787‐7 Dimethoate Cheminova Inc [Third Party: Lewis & Harrison LLC] Warning Cheminova Dimethoate 4E 34704‐207‐67760 Dimethoate Cheminova Inc [Third Party: Lewis & Harrison LLC] Warning Cheminova Methyl 4EC 67760‐43 Methyl parathion Cheminova Inc [Third Party: Lewis & Harrison LLC] Danger/Poison Chlorpyrifos 4E AG 66222‐19 Chlorpyrifos Makhteshim‐Agan of North America Inc Warning Chlorpyrifos 4E AG 66330‐278 Chlorpyrifos Arysta LifeScience North America, LLC Warning Cobalt Insecticide 62719‐575 Chlorpyrifos, Gamma‐ Dow AgroSciences Inc Danger cyhalothrin Counter 15G Lock'n Load 5481‐545 Terbufos Amvac Chemical Corporation Danger/Poison Counter 15G Smartbox Systemic 5481‐545 Terbufos Amvac Chemical Corporation Danger/Poison