Microfossil Assemblages, Zonations and Planktonic Datum Levels in the Ieper Formation (Ypresian S .S ., Early Eocene) in Belgium
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

73 Lijn 73 : Gent - Deinze - De Panne
nmbs 73 Lijn 73 : Gent - Deinze - De Panne Stations en haltes • AGENT-SINT-PIETERS • YDE PINTE • YDEINZE • YAARSELE • YTIELT • YLICHTERVELDE • YKORTEMARK • YDIKSMUIDE • YVEURNE • YKOKSIJDE • CDE PANNE Dienstregelingen geldig vanaf 13.12.2020 Uitgave 13.06.2021 Conventionele tekens ì Rijdt tijdens de jaarlijkse vakantieperiodes 19.12.20 → 02.01.21 03.04.21 → 17.04.21 03.07.21 → 29.08.21 í Rijdt niet tijdens de jaarlijkse vakantieperiodes Rijdt tijdens de toeristische periode (zomer) èí 03.07.21 → 29.08.21 3 Op weekdagen, behalve feestdagen 4 Op zaterdagdagen, zon- en feestdagen ,...2 Maandag...Zondag ^ Zon- en feestdagen + Aankomstuur van de trein [ Diabolotoeslag van toepassing voor deze bestemming @ InterCitytrein ` Voorstadstrein = Lokale trein > Piekuurtrein ç Bus å Toeristische trein Ü Thalys trein à ICE trein (Deutsche Bahn) Feestdagen gelijkgesteld met een zondag : 25.12.20 - 01.01.21 - 05.04.21 - 01.05.21 - 13.05.21 - 24.05.21 - 21.07.21 - 01.11.21 - 11.11.21 Brugdagen waarop de treindienst kan worden aangepast : Op 14.05.21 en 12.11.21 Voor de laatste updates : Plan uw reis via de NMBS-app of nmbs.be Maandag tot vrijdag, behalve op feestdagen 73 Gent - Deinze - De Panne @ @ @ å @ @ @ @ 3004 3005 3006 6901 3007 3008 3009 3010 è Herkomst Antw. Antw. Antw. Antw. Antw. Cent. Cent. Cent. Cent. Cent. 6.50 7.51 8.51 9.51 10.51 Brus.-Noord 7.27 Brus.-Centraal 7.32 Brus.-Zuid 7.45 Gent-Sint-Pieters + 8.14 Gent-Sint-Pieters 5.53 6.53 7.53 8.21 8.53 9.53 10.53 11.53 De Pinte + 6.00 7.00 8.00 8.28 9.00 10.00 11.00 12.00 De Pinte 6.01 7.01 -

Adressen Diensten Voor Oppashulp West-Vlaanderen Repertorium Diensten Voor Oppashulp West-Vlaanderen
Agentschap Zorg en Gezondheid Afdeling Preventie, Eerstelijn en Thuiszorg Koning Albert II-laan 35 bus 33, 1030 Brussel Tel. 02-553 36 47 - Fax 02 553 36 05 Adressen Diensten voor Oppashulp West-Vlaanderen Repertorium Diensten voor Oppashulp West-Vlaanderen Voorziening Beheersinstantie BRUGGE Familiezorg West-Vlaanderen Regio Brugge- Familiezorg West-Vlaanderen Oostende Dossiernr.: 502 VZW Erkenningsnr.: OPP502 Sint-Jansplein 8 Biskajersplein 3 8000 Brugge 8000 Brugge tel. : fax: website: e-mail: Oppas Regio Brugge-Oostende Oppas Dossiernr.: 522 VZW Erkenningsnr.: OPP522 Oude Burg 27 Oude Burg 27 8000 Brugge 8000 Brugge tel. : fax: website: e-mail: Pagina 2 van 9 6-sep-21 Agentschap Zorg en Gezondheid Afdeling Woonzorg en Eerste Lijn Koning Albert II-laan 35 bus 33, 1030 Brussel Tel. 02-553 36 47 - Fax 02 553 36 05 Repertorium Diensten voor Oppashulp West-Vlaanderen Voorziening Beheersinstantie IEPER Familiezorg West-Vlaanderen Regio Veurne- Familiezorg West-Vlaanderen Diksmuide-Ieper Dossiernr.: 503 VZW Erkenningsnr.: OPP503 De Brouwerstraat 4 Biskajersplein 3 8900 Ieper 8000 Brugge tel. : fax: website: e-mail: Pagina 3 van 9 6-sep-21 Agentschap Zorg en Gezondheid Afdeling Woonzorg en Eerste Lijn Koning Albert II-laan 35 bus 33, 1030 Brussel Tel. 02-553 36 47 - Fax 02 553 36 05 Repertorium Diensten voor Oppashulp West-Vlaanderen Voorziening Beheersinstantie KORTRIJK Oppas Zuid-West-Vlaanderen OPPAS ZUID-WEST-VLAANDEREN Dossiernr.: 513 VZW Erkenningsnr.: OPP513 Beneluxpark 22 Beneluxpark(Kor) 22 8500 Kortrijk 8500 Kortrijk tel. : fax: website: e-mail: i-mens Regio Kortrijk-Roeselare-Tielt i-mens Dossiernr.: 526 VZW Erkenningsnr.: OPP526 't Hoge 49 Tramstraat 61 8500 Kortrijk 9052 Gent tel. -

Provincieraadsleden Legislatuur 2018-2024
Provincieraadsleden legislatuur 2018-2024 Naam Voornaam Fractie District Gemeente Aernoudt Wim N-VA Ieper - Oostende - Diksmuide Gistel Bekaert Simon sp.a Kortrijk - Roeselare - Tielt Tielt Coupillie Luc N-VA Ieper - Oostende - Diksmuide Diksmuide Croes Claude CD&V Kortrijk - Roeselare - Tielt Deerlijk de Bethune Jean CD&V Kortrijk - Roeselare - Tielt Kortrijk De Klerck Patrick Open Vld Brugge Blankenberge De Poorter Johan N-VA Kortrijk - Roeselare - Tielt Meulebeke De Reuse Immanuel Vlaams Belang Kortrijk - Roeselare - Tielt Roeselare Decleer Hilde CD&V Brugge Brugge Dejaegher Christof CD&V Ieper - Oostende - Diksmuide Poperinge Dejaeghere Veerle Groen Kortrijk - Roeselare - Tielt Ardooie Dumarey Anthony Open Vld Ieper - Oostende - Diksmuide Oudenburg Hollevoet Justine sp.a Kortrijk - Roeselare - Tielt Izegem Kindt Els CD&V Kortrijk - Roeselare - Tielt Lichtervelde Lahaye-Battheu Sabien Open Vld Ieper - Oostende - Diksmuide Poperinge Laridon Lies CD&V Ieper - Oostende - Diksmuide Diksmuide Lodewyckx Herman Groen Ieper - Oostende - Diksmuide Oostende Moerman Bieke sp.a Ieper - Oostende - Diksmuide Diksmuide Naeyaert Bart CD&V Brugge Torhout Pillaert Kristof N-VA Kortrijk - Roeselare - Tielt Hooglede Ravyts Kurt Vlaams Belang Ieper - Oostende - Diksmuide Zuienkerke Schotte Gerda Groen Brugge St. Michiels Sneppe Dominiek Vlaams Belang Brugge Zedelgem Soens Rik CD&V Kortrijk - Roeselare - Tielt Beveren(Leie) Strobbe Ruben CD&V Brugge Beernem Tavernier Maarten Groen Kortrijk - Roeselare - Tielt Wevelgem Van Meirhaeghe Jan N-VA Brugge Damme Vanbrabant Martine Vlaams Belang Ieper - Oostende - Diksmuide Poperinge Vandenbrande Isabelle N-VA Brugge Beernem Vandenbulcke Sigrid N-VA Kortrijk - Roeselare - Tielt Menen Vandermersch Piet Open Vld Kortrijk - Roeselare - Tielt Ledegem Vanlerberghe Jurgen Spa Ieper - Oostende - Diksmuide Poperinge Vannieuwenhuyze Luc CD&V Kortrijk - Roeselare - Tielt Tielt Vanryckeghem Martine Open Vld Kortrijk - Roeselare - Tielt Rekkem Vermeersch Wouter Vlaams Belang Kortrijk - Roeselare - Tielt Kortrijk Willems Tom sp.a Brugge Brugge . -
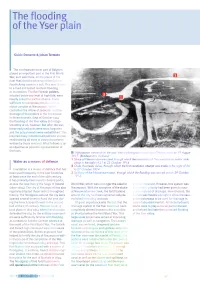
The Flooding Ofthe Yser Plain
The flooding ofthe Yser plain Guido Demerre & Johan Termote The northwesternmost part of Belgium played an important part in the First World War, as it was there, on the plains of the river Yser, that the advance of the German Fourth Army came to a halt. This was thanks to a tried and tested method: flooding or inundation. The flat Flemish polders, situated below sea level at high tide, were ideally suited for such a scheme. It was sufficient to manipulate the Ganzepoot sluice complex at Nieuwpoort, which controlled the inflow of seawater and the drainage ofthe polders in the hinterland. In those dramatic days of October 1914 the flooding of the Yser valley did not go smoothly at all, however. But after the war, these early setbacks were soon forgotten and the actual events were embellished. This inspired many romanticised patriotic stories, reinforced by all sorts of vanity documents written by those involved. What follows is an as objective as possible representation of the facts. Nieuwpoort viewed from the east. Aerial photograph made by a German scout on 19 August 1917. (Bundesarchiv, Koblenzj 1 Sluice at Nieuwendamme creek through which the inundation of Nieuwendamme polder took Water as a means of defence place in the night o f 21 to 22 October 1914 2 Oude Veurnesas sluice, through which the first inundation attempt was made in the night o f 26 Inundation is a means of defence that has to 27 October 1914 been used frequently in the Low Countries 3 Spillway of the Noordvaart canal, through which the flooding was carried out on 29 October at least since the end of the 16th century. -

Curriculum Vitae
CURRICULUM VITAE STEVEN PETERS CARABALLO EDUCATION 1997 – 2001 Master Degree in Graphical Design (Academy of Fine Arts, Genk, Belgium) RESIDENCIES 2019 Arteventura (27/02 – 16/05) Aracena (SP) EXHIBITIONS Upcoming Gallery De Wael, April 2020, Antwerp, Belgium CAS (Contemporary Art Space) July 2020, Zottegem, Belgium A Butterfly’s Scream, September 2020, Beernem, Belgium Gallery De Wael, Solo show, September 2020, Beernen, Belgium 2020 Syndicate of Belgian Painters (11/01 – 08/03), St Truiden (Belgium), Borgloon (Belgium), Diepenbeek (Belgium) 2019 Re-Search (05/09 – 26/10) Riboli Building Brussels, Belgium Come Closer (13/07 – 29/09) Rekem, Belgium Salon Blanc (15/06) Oostende, Belgium Anouk Vilain Gallery (01/06 – 30/06) Diepenbeek, Belgium Model Staan (03/05 – 01/06) Tielt, Belgium Art-fair @ Quartier Bleu (17/03 – 24/03) Hasselt, Belgium 2018 Dark Rooms (29/10 - 17/12) PAK, Distel, Belgium Art at the Villa (12/10 – 26/08) Kraainem, Belgium Bienale ART-AUTUN, Ce qui pese et ce qui nourrit (07/07 – 26/08) Autun, France CAS (Contemporary Art Space) Kunstroute (17/06 - 01/07) Zottegem, Belgium Behind Closed Doors (16/06 – 15/07) PAK Gistel, Belgium Faces (10/03 – 26/04) Sint-Amandus, Belgium 2017 CAS (Contemporary Art Space) (11/11 – 26/11) Zottegem, Belgium Arture (15/09 – 16/09) Herzele, Belgium Arture (03/06 – 10/06) Herzele, Belgium Inner Mind (129/05 – 30/06) De Haan, Belgium Between Heaven and Earth (23/04 – 25/06) Brugge, Belgium 2016 I Presume (17/11 – 20/11) Dak, Utrecht, Netherlands Drawing Rooms 2 (11/09 – 16/10) PAK Gistel, Belgium The Protagonist (02/04 – 24/04) Wellen, Belgium 2015 Ronse Drawing Prize (Selection) (05/12 – 31/12) De Ververii, Ronse, Belgium Actueel Kunsten Festival (31/10 – 19/11) Heks, Heers, Kuttekove, Wellen, Belgium Text Through Steven Peters Caraballo’s works, classic themes such as the portrait, the still life and the vanitas are again given a chance to distinguish themselves through the magnificence of their simplicity. -

PDF Immobel Belgium Press Release Tielt EN
PRESS RELEASE 22/06/2021 Non-regulated information Brussels, 22 June at 11 AM CET GREEN LIGHT FOR A GREEN LIVING OASIS IN TIELT Immobel and Dumobil have obtained planning permission for the development of a completely new, green area in the West Flanders town of Tielt. Both the town and provincial councils have approved the parcelling of this ambitious housing project. Sustainable housing units will be combined harmoniously with the creation of a vast new park. The start of the work is planned for the autumn of 2021. Almost 200 units to be built in a park of 48,000m² Project developer Immobel will draw on more than two centuries of experience for this project. Specifically, Immobel will create a park surrounded by bespoke homes. There will be a mixture of 106 houses and 88 apartments. This means that the town of Tielt will see its population increase by around 1,000 inhabitants over time. The various living units will answer to the strictest insulation and energy norms and blend harmoniously with the surrounding countryside. The new park, with numerous colourful wildflower beds and more than 300 oak, linden and other indigenous trees, will promote relaxation and neighbourly encounters. Alongside sustainability, mobility also holds an important role. With its central location, close to the town centre and Tielt railway station, the project represents a nice upgrade for the urban environment. Green living “The park, which covers almost 5 hectares, forms the heart of this unique living project.” says Toon Honings, Development Director Immobel Home. “This project is an example of our philosophy of sustainable living. -

Regional Development Programmes Belgium 1978-1980
COMMISSION OF THE EUROPEAN COMMUNITIES programmes Regional development programmes Belgium 1978-1980 REGIONAL POLICY SERIES - 1979 14 COMMISSION OF THE EUROPEAN COMMUNITIES Regional development programmes Belgium 1978-1980 COLLECTION PROGRAMMES Regional Policy Series No 14 Brussels, November 1978 NOTE It is appropriate first of all to draw the reader's attention to the essentially indicative character of these regional development programmes. This publication is also available in DE ISBN 92-825-0960-5 FR ISBN 92-825-0962-1 NL ISBN 92-825-0963-X Cataloguing data can be found at the end of this volume © Copyright ECSC - EEC - EAEC, Brussels - Luxembourg, 1979 Reproduction authorized, in whole or in part, provided the source is acknowledged. Printed in Luxembourg ISBN 92-825-0961-3 Catalogue number: CB-NS-79-014-EN-C Contents GENERAL INTRODUCTION 5 Detailed index 7 A. Socio-economic development trends in Belgium, 1960-1977 9 B. National political options 11 C. The regional development programmes 12 D. The constitutional institutions 15 Annexes 21 Maps 33 REGIONAL DEVELOPMENT PROGRAMME FLANDERS 1978-1980 37 Detailed index 39 Chapter I - Economic and social analysis 43 Chapter II - Development objectives 91 Chapter III - Development measures 105 Chapter IV - Financial resources 129 Chapter V - Implementation of the programme 139 Annex 145 REGIONAL DEVELOPMENT PROGRAMME WALLONIA 1978-1980 147 Detailed index 149 Chapter I - Economic and social analysis 151 Chapter II - Development objectives 189 Chapter III - Development measures 193 Chapter IV - Financial resources 201 Chapter V - Implementation of the programme 207 Annexes 213 General introduction Detailed index A. Socio-economic development trends in Belgium, 1960-1977 9 B. -

Communication to the Commission On
EN EN EN COMMISSION OF THE EUROPEAN COMMUNITIES Brussels, xxx SEC(2009) 1154 final COMMUNICATION TO THE COMMISSION on applications EGF/2009/004 BE/Oost en West Vlaanderen textiles and EGF/2009/005 BE/Limburg textiles received from Belgium for a financial contribution from the European Globalisation Adjustment Fund EN EN COMMUNICATION TO THE COMMISSION on applications EGF/2009/004 BE/Oost en West Vlaanderen textiles and EGF/2009/005 BE/Limburg textiles received from Belgium for a financial contribution from the European Globalisation Adjustment Fund Belgium has submitted applications EGF/2009/004 BE/Oost en West Vlaanderen textiles and EGF/2009/005 BE/Limburg textiles for a financial contribution from the European Globalisation Adjustment Fund (hereinafter 'EGF') following redundancies in enterprises operating in NACE Revision 2 division 13 ('manufacture of textiles') in two contiguous NUTS II regions of East (BE230) and West Flanders (BE225) and a single NUTS II region, Limburg (BE220), respectively. 1. The two applications were presented to the Commission on 5 May 2009 and supplemented by additional information on 29 June 2009. 2. In accordance with Article 2 of Regulation (EC) N°546/2009 of the European Parliament and of the Council of 18 June 2009 amending Regulation (EC) N°1927/2006 on establishing the European Globalisation Adjustment Fund1, the applications were assessed on the basis of Regulation (EC) N°1927/2006 of the European Parliament and of the Council of 20 December 2006 on establishing the European Globalisation Adjustment Fund2 as amended by Regulation (EC) N°546/2009. 3. The two applications meet the conditions for deploying the EGF as set out in Article 2(b) of Regulation (EC) N°1927/2006, and were submitted within the deadline of 10 weeks provided for in Article 5 of that Regulation. -

Ondervoorzitter Van De Vrederechters En Rechters in De Politierechtbank Van Het Arrondissement West-Vlaanderen
Ondervoorzitter van de vrederechters en rechters in de politierechtbank van het arrondissement West-Vlaanderen Rep. nr. 2020/59 BESCHIKKING TOT VASTSTELLING VAN HET BIJZONDER REGLEMENT VAN DE VREDEGERECHTEN WEST-VLAANDEREN Rekening houdend met: de artikelen 66, 335bis, 670 en 1035 van het gerechtelijk wetboek, de wet van 15 juni 1935 op het gebruik der talen in gerechtszaken, artikel 43§2 van het Vlaams Woninghuurdecreet, het advies van de stafhouder van de Orde van Advocaten te West-Vlaanderen, verleent de ondervoorzitter van de vrederechters en de rechters in de politierechtbank van het arrondissement West-Vlaanderen, bijgestaan door de hoofdgriffier, de volgende beschikking: 1. De vredegerechten over het grondgebied van het gerechtelijk arrondissement West- Vlaanderen bestaan uit 17 kantons: - vredegerecht eerste kanton Brugge - vredegerecht tweede kanton Brugge - vredegerecht derde kanton Brugge - vredegerecht vierde kanton Brugge - vredegerecht kanton Ieper - vredegerecht kanton Izegem - vredegerecht eerste kanton Kortrijk - vredegerecht tweede kanton Kortrijk - vredegerecht kanton Menen - vredegerecht eerste kanton Oostende - vredegerecht tweede kanton Oostende - vredegerecht kanton Poperinge - vredegerecht kanton Roeselare - vredegerecht kanton Tielt - vredegerecht kanton Torhout - vredegerecht kanton Veurne - vredegerecht kanton Waregem 2. Het aantal en de dagen van de gewone openbare zittingen van de vredegerechten van het arrondissement West-Vlaanderen voor de periode van 1 september tot 30 juni (gerechtelijk jaar) worden -

Business Centres in the Province of West-Flanders
KNO(!KKE-HEIST BLANKENBERGE Business Centre 't Walletje Baudouin Canal Business Centre Ostend (OC) E34 BEVEREN Bluebridge - Ostend Science Park (! (! OSTEND (! Business Centre Ostend - Zandvoordestraat (! (! (!(! (!BRUGES A10 E4(!03(!(!(!(! (! E34 (! EE(!KLO Vaartdijk Business Centre Business Centre Veurne - Acasus (! E40 Business Centre Ostend - Kuipweg Canal Ghent-Ostend (! (! (!TEMSE Business Centre TVELT E40 Office 407 (! BLACKBOXX (! ! (! Business Centre 3koningen ((! (! IC Grote Beer Business Centre Bruges (! LOKCEORERNE BRUGES (! (! Business Centre Veurne (! (! Bus(!iness Centre Bruges (OC) (! (! E17 TORHOUT E40 (! (! (! (!(!(!(! (!(! Business Centre Diksmuide E403 (!(!(! (! (! Business Centre Wingene (!(!(!(!(! BRUGGE.INC VEURNE DIKSMUIDE Business Centre Wingene (Driebeck) (!W(!ork-Around (! (!(! (! (! (! (! DENDERMONDE DEINZE (! (!(!(!(! (! (! IJzer T(!IELT (!(! WETTERE(!N Business Centre Roeselare REGUS Bruges Express (! Katelijnepoort Business Centre (! Vlaemynck Business Cen(!tre (! Vaartdijk Business Centre (! ROESELARE (! The Vibe Business Centre Izegem (! Business Centre Roeselare (OC) E17 (! (!(! Atelier12 (! REGUS Roeselare West Wing Park ! (! Leie (!((! Business Centre Poperinge d'Office Business Centre Ieper E40 ASSE Business Centre Waregem (! ZOTTEGEM (! (! WA(!REG(!EM (! Stede 51 (! POPERIN(!GE IEPER (! OUDENAARDE (! A19 (! E17 Stede 51 ! (!(!(! (! CORE KORTRIJK (! (!(!((! (! ABC-Waregem (! (! Business Centre Waregem (Region) Studio Ko - Coworking Kortrijk MENEN Leie KORTRIJK Hangar K Business Centre Kortrijk (! Start -

Vervoerregio Loopnummerpubliek Lijnnummer Bestemming Rit Rit
Vervoerregio LoopnummerPubliek lijnnummer Bestemming rit Rit Startplaats Startuur einduur eindplaats Maandag Dinsdag Woensdag Donderdag Vrijdag Kortrijk 4 40 Menen 51 Kortrijk Busterminal 2 07:22 07:53 Menen Station x x x x x Kortrijk 4 40 kortrijk 150 Menen Station 12:25 12:57 Kortijk Busterminal 2 x Kortrijk 4 40 kortrijk 230 Menen Station 16:11 16:49 Kortijk Busterminal 2 x x x x Kortrijk 5 62 Roeselare 21 Kortrtijk Busterminal 2 07:03 08:01 Roeselare Station x x x x x Kortrijk 5 62 kortrijk 40 Roeselare Station 12:22 13:12 Kortrtijk Busterminal 2 x Kortrijk 5 62 kortrijk 70 Roeselare Station 16:28 17:20 Kortrtijk Busterminal 2 x x x x Kortrijk 1 72 Kortrijk Busterminal4 20 Tiegem Plaats 07:07 08:02 Kortrijk Busterminal 4 x x x x x Kortrijk 1 13 Kortrijk Universtiteit 81 Kortrijk Busterminal 3 08:08 08:23 Kortrijk Universiteit x x x x x Kortrijk 1 13 Kortrijk Busterminal 3 80 Kortrijk Universiteit 08:23 08:27 Kortrijk Busterminal 3 x x x x x Kortrijk 1 13 Kortrijk Universtiteit 171 Kortrijk Busterminal 3 08:40 08:52 Kortrijk Universiteit x x x x x Kortrijk 1 13 Kortrijk Busterminal 3 170 Kortrijk Universiteit 08:52 08:56 Kortrijk Busterminal 3 x x x x x Kortrijk 1 13 Kortrijk Busterminal 3 480 Kortrijk Universiteit 16:17 16:32 Kortrijk Busterminal 3 x x x x Kortrijk 1 13 Kortrijk Busterminal 3 530 Kortrijk Universiteit 17:03 17:18 Kortrijk Busterminal 3 x x x x Kortrijk 8 75 Deinze 60 Kortrijk Busterminal 4 06:47 08:01 Deinze Station x x x x x Kortrijk 8 75 Desselgem 211 Deinze Kerkhof 12:10 12:58 Desselgem Toren x Kortrijk 8 75 Desselgem -

Voorstelling Statistieken 2017
2017 Jaarverslag openbaredienstverplichting 1 1 Jaarverslag 2017 Overlegorgaan openbaredienstverplichting tot het vervoer van personen met een handicap of een ernstig beperkte mobiliteit 2 2 Statistieken vervoer Ritten Niet uitgevoerde ritten Klachten & doorverwijzingen Software & internetgebruik 3 3 Ritinformatie : Aantal aangevraagde ritten Aantal uitgevoerde ritten en % rolstoelgebruikers Aantal niet uitgevoerde ritten 4 4 Ritten 2017 Aangevraagd (100%) 291.821 Uitgevoerde ritten Uitgevoerd (85%) 248.051 Rolstoel (65%) 236.151 5 5 Evolutie uitgevoerde ritten Cijfers uitgevoerde ritten aantal uitvoering 2014: 193.534 84% 2015: 205.930 86% 2016: 223.439 86% 2017: 248.051 86% Procentuele groei vanaf 2014 2015 : + 6 % 2016 : + 9 % 2017 : + 11 % 6 6 rolstoelgebruikers 2017 : 236.151 ( gemiddeld 65%) Spreiding : 10 vervoerders meer dan 75% rolstoelklanten 8 vervoerders tussen 50% en 75% rolstoelklanten 7 vervoerders tussen 25% en 50 % rolstoelklanten 2 vervoerders minder dan 25% rolstoelklanten 7 7 Evolutie spreiding vervoerders met % rolstoelklanten +75 ts 50 en 75 ts 25 en 50 ts 0 en 25 vervoerders 2014 15 5 2 2 24 2015 6 6 3 2 17 2016 11 7 6 1 25 2017 10 8 7 2 27 8 8 2.139.872 km gereden in 2017, gereden door : 5 vervoerders meer dan 150.000 km per jaar 4 vervoerders tussen 100.000 km en 150.000 km per jaar 5 vervoerders tussen 50.000 km en 100.000 km per jaar 8 vervoerders tussen 25.000 km en 50.000 km per jaar 5 vervoerders minder dan 25.000 km per jaar 9 9 Evolutie spreiding gereden kilometers 2014 2015 2016 2017 meer dan 150 3 3 4