Unpacking the Species Conundrum: Philosophy, Practice and a Way Forward
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Of the Scinax Ruber Clade from Cerrado of Central Brazil
Amphibia-Reptilia 31 (2010): 411-418 A new species of small Scinax Wagler, 1830 (Amphibia, Anura, Hylidae) of the Scinax ruber clade from Cerrado of central Brazil Manoela Woitovicz Cardoso, José P. Pombal Jr. Abstract. A new species of the Scinax ruber clade from the Brazilian Cerrado Domain similar to Scinax fuscomarginatus, S. parkeri, S. trilineatus and S. wandae is described. It is characterized by small snout-vent lenght, body slender, head approximately as long as wide and slightly wider than body, subovoid snout in dorsal view, protruding snout in lateral view, a developed supratympanic fold, absence of flash colour on the posterior surfaces of thighs, hidden portions of shanks and groin, and large vocal sac. Scinax lutzorum sp. nov. differs from S. fuscomarginatus, S. parkeri and S. trilineatus by its slightly larger SVL; from Scinax fuscomarginatus and S. parkeri it differs by its more slender body; from Scinax fuscomarginatus and S. trilineatus the new species differs by its wider head and more protruding eyes; and it differs from Scinax parkeri and S. wandae by its shorter snout. Comments on the type specimens of S. fuscomarginatus are presented and a lectotype is designated for this species. Keywords: lectotype, new species, Scinax fuscomarginatus, Scinax lutzorum. Introduction 1862), S. cabralensis Drummond, Baêta and Pires, 2007, S. camposseabrai (Bokermann, The hylid frog genus Scinax Wagler, 1830 cur- 1968), Scinax castroviejoi De La Riva, 1993, rently comprises 97 recognized species distrib- S. curicica Pugliese, Pombal and Sazima, 2004, uted from eastern and southern Mexico to Ar- S. eurydice (Bokermann, 1968), S. fuscomar- gentina and Uruguay, Trinidad and Tobago, and ginatus (A. -

North Andean Origin and Diversification of the Largest Ithomiine Butterfly Genus
North Andean origin and diversification of the largest ithomiine butterfly genus The Harvard community has made this article openly available. Please share how this access benefits you. Your story matters Citation Lisa De-Silva, D., L. L. Mota, N. Chazot, R. Mallarino, K. L. Silva- Brandão, L. M. G. Piñerez, A. V. Freitas, et al. 2017. “North Andean origin and diversification of the largest ithomiine butterfly genus.” Scientific Reports 7 (1): 45966. doi:10.1038/srep45966. http:// dx.doi.org/10.1038/srep45966. Published Version doi:10.1038/srep45966 Citable link http://nrs.harvard.edu/urn-3:HUL.InstRepos:32630680 Terms of Use This article was downloaded from Harvard University’s DASH repository, and is made available under the terms and conditions applicable to Other Posted Material, as set forth at http:// nrs.harvard.edu/urn-3:HUL.InstRepos:dash.current.terms-of- use#LAA www.nature.com/scientificreports OPEN North Andean origin and diversification of the largest ithomiine butterfly genus Received: 31 October 2016 Donna Lisa De-Silva1, Luísa L. Mota2, Nicolas Chazot1,3, Ricardo Mallarino4, Karina L. Silva- Accepted: 22 February 2017 Brandão5, Luz Miryam Gómez Piñerez6,7, André V.L. Freitas2, Gerardo Lamas8, Published: 07 April 2017 Mathieu Joron9, James Mallet4, Carlos E. Giraldo6,10, Sandra Uribe6, Tiina Särkinen11, Sandra Knapp12, Chris D. Jiggins13, Keith R. Willmott14 & Marianne Elias1 The Neotropics harbour the most diverse flora and fauna on Earth. The Andes are a major centre of diversification and source of diversity for adjacent areas in plants and vertebrates, but studies on insects remain scarce, even though they constitute the largest fraction of terrestrial biodiversity. -

Identification of the Taxonomic Status of Scinax Nebulosus and Scinax Constrictus (Scinaxinae, Anura) Based on Molecular Markers T
Brazilian Journal of Biology https://doi.org/10.1590/1519-6984.225646 ISSN 1519-6984 (Print) Original Article ISSN 1678-4375 (Online) Identification of the taxonomic status of Scinax nebulosus and Scinax constrictus (Scinaxinae, Anura) based on molecular markers T. M. B. Freitasa* , J. B. L. Salesb , I. Sampaioc , N. M. Piorskia and L. N. Weberd aUniversidade Federal do Maranhão – UFMA, Departamento de Biologia, Laboratório de Ecologia e Sistemática de Peixes, Programa de Pós-graduação Bionorte, Grupo de Taxonomia, Biogeografia, Ecologia e Conservação de Peixes do Maranhão, São Luís, MA, Brasil bUniversidade Federal do Pará – UFPA, Centro de Estudos Avançados da Biodiversidade – CEABIO, Programa de Pós-graduação em Ecologia Aquática e Pesca – PPGEAP, Grupo de Investigação Biológica Integrada – GIBI, Belém, PA, Brasil cUniversidade Federal do Pará – UFPA, Instituto de Estudos Costeiros – IECOS, Laboratório e Filogenomica e Bioinformatica, Programa de Pós-graduação Biologia Ambiental – PPBA, Grupo de Estudos em Genética e Filogenômica, Bragança, PA, Brasil dUniversidade Federal do Sul da Bahia – UFSB, Centro de Formação em Ciências Ambientais, Instituto Sosígenes Costa de Humanidades, Artes e Ciências, Departamento de Ciências Biológicas, Laboratório de Zoologia, Programa de Pós-graduação Bionorte, Grupo Biodiversidade da Fauna do Sul da Bahia, Porto Seguro, BA, Brasil *e-mail: [email protected] Received: June 26, 2019 – Accepted: May 4, 2020 – Distributed: November 30, 2021 (With 4 figures) Abstract The validation of many anuran species is based on a strictly descriptive, morphological analysis of a small number of specimens with a limited geographic distribution. The Scinax Wagler, 1830 genus is a controversial group with many doubtful taxa and taxonomic uncertainties, due a high number of cryptic species. -

Butterflies (Lepidoptera: Papilionoidea) in a Coastal Plain Area in the State of Paraná, Brazil
62 TROP. LEPID. RES., 26(2): 62-67, 2016 LEVISKI ET AL.: Butterflies in Paraná Butterflies (Lepidoptera: Papilionoidea) in a coastal plain area in the state of Paraná, Brazil Gabriela Lourenço Leviski¹*, Luziany Queiroz-Santos¹, Ricardo Russo Siewert¹, Lucy Mila Garcia Salik¹, Mirna Martins Casagrande¹ and Olaf Hermann Hendrik Mielke¹ ¹ Laboratório de Estudos de Lepidoptera Neotropical, Departamento de Zoologia, Universidade Federal do Paraná, Caixa Postal 19.020, 81.531-980, Curitiba, Paraná, Brazil Corresponding author: E-mail: [email protected]٭ Abstract: The coastal plain environments of southern Brazil are neglected and poorly represented in Conservation Units. In view of the importance of sampling these areas, the present study conducted the first butterfly inventory of a coastal area in the state of Paraná. Samples were taken in the Floresta Estadual do Palmito, from February 2014 through January 2015, using insect nets and traps for fruit-feeding butterfly species. A total of 200 species were recorded, in the families Hesperiidae (77), Nymphalidae (73), Riodinidae (20), Lycaenidae (19), Pieridae (7) and Papilionidae (4). Particularly notable records included the rare and vulnerable Pseudotinea hemis (Schaus, 1927), representing the lowest elevation record for this species, and Temenis huebneri korallion Fruhstorfer, 1912, a new record for Paraná. These results reinforce the need to direct sampling efforts to poorly inventoried areas, to increase knowledge of the distribution and occurrence patterns of butterflies in Brazil. Key words: Atlantic Forest, Biodiversity, conservation, inventory, species richness. INTRODUCTION the importance of inventories to knowledge of the fauna and its conservation, the present study inventoried the species of Faunal inventories are important for providing knowledge butterflies of the Floresta Estadual do Palmito. -

Revealing Cryptic Diversity Using Molecular Phylogenetics and Phylogeography in Frogs of the Scinax Ruber and Rhinella Margaritifera Species Groups
Molecular Phylogenetics and Evolution 43 (2007) 567–582 www.elsevier.com/locate/ympev Revealing cryptic diversity using molecular phylogenetics and phylogeography in frogs of the Scinax ruber and Rhinella margaritifera species groups Antoine Fouquet a,f,¤, Miguel Vences b, Marie-Dominique Salducci a, Axel Meyer c, Christian Marty d, Michel Blanc e, André Gilles a a EA 3781 EGEE Evolution Genome Environment, Université de Provence, Centre St. Charles, 3 place Victor Hugo, 13331 Marseille, France b Zoological Institute, Technical University of Braunschweig, Spielmannstr. 8, 38106 Braunschweig, Germany c Department of Biology, Evolutionary Biology, University of Konstanz, 78457 Konstanz, Germany d Impasse Jean Galot, 97354 Montjoly, Guyane Française e 2 rue Doct. Floch, 97310 Kourou, Guyane Française f Molecular Ecology Laboratory, School of Biological Sciences, University of Canterbury, Private Bag 4800, Christchurch, New Zealand Received 25 April 2006; revised 27 November 2006; accepted 1 December 2006 Available online 23 December 2006 Abstract Few studies to date have examined genetic variability of widespread tropical amphibian species over their distributional range using diVer- ent kinds of molecular markers. Here, we use genetic data in an attempt to delimit evolutionary entities within two groups of Neotropical frogs, the Scinax ruber species group and the Rhinella margaritifera species group. We combined mitochondrial and nuclear markers for a phylogenetic (a total of »2500 bp) and phylogeographic study (»1300 bp) to test the reliability of the currently accepted taxonomic assign- ments and to explore the geographic structure of their genetic variation, mainly based upon samples from the French Guianan region. Phylo- genetic analyses demonstrated the polyphyly of Scinax ruber and Rhinella margaritifera. -

Calls of Five Species of the Scinax Ruber (Anura: Hylidae) Clade from Brazil with Comments on Their Taxonomy
Zootaxa 3066: 37–51 (2011) ISSN 1175-5326 (print edition) www.mapress.com/zootaxa/ Article ZOOTAXA Copyright © 2011 · Magnolia Press ISSN 1175-5334 (online edition) Calls of five species of the Scinax ruber (Anura: Hylidae) clade from Brazil with comments on their taxonomy LEANDRO MAGRINI1,2,5, SERGIO P. DE CARVALHO-E-SILVA3, ARLINDO F. BÉDA4 & ARIOVALDO A. GIARETTA2 1Departamento de Biologia - Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto. Universidade de São Paulo, Ribeirão Preto, São Paulo, Brazil. CEP 14040-901. 2Laboratório de Taxonomia e Sistemática de Anuros Neotropicais. Facip. Universidade Federal de Uberlândia, Ituiutaba, Minas Gerais, Brazil. CEP: 38302–000. E-mail: [email protected] 3Laboratório de Anfíbios e Répteis, Departamento de Zoologia, Instituto de Biologia, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Rio de Janeiro, Brazil. CEP: 68044. 4Campus de Aquidauana, Universidade Federal de Mato Grosso do Sul, Aquidauana, Mato Grosso do Sul, Brazil. CEP: 79200-000. 5Corresponding author. E-mail: [email protected] Abstract As currently defined, tree frogs of the genus Scinax comprises the most species-rich genus within Hylinae. Although in the last decades there have been an increasing number of taxonomic studies on Scinax, populations of several species still deserve further studies to assess their taxonomic status. The purpose of this paper is to contribute to the taxonomy and zoogeography of some species of Scinax from Brazil assigned to the Scinax ruber clade through the description/redescrip- tion of their advertisement calls. The advertisement call of S. duartei from topotypic specimens and the call of S. acumi- natus are described here for the first time. -
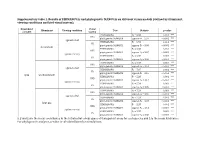
Supplementary Table 1. Results of Permanovas and Phylogenetic Manovas on Different Vision Models (Defined by Illuminant, Viewing Conditions and Bird Visual System)
Supplementary table 1. Results of PERMANOVAs and phylogenetic MANOVAs on different vision models (defined by illuminant, viewing conditions and bird visual system). Dependent Visual Illuminant Viewing condition Test Statistic p-value variable system PERMANOVA F9 = 6.88 0.001 *** UVS phylogenetic MANOVA approx-F9 = 2.97 < 0.001 *** against a leaf PERMANOVA F9 = 6.93 0.001 *** VS phylogenetic MANOVA approx-F9 = 3.05 < 0.001 *** forest shade PERMANOVA F9 = 5.38 0.001 *** UVS phylogenetic MANOVA approx-F9 = 3.07 < 0.001 *** against the sky PERMANOVA F9 = 5.38 0.001 *** VS phylogenetic MANOVA approx-F9 = 3.36 < 0.001 *** PERMANOVA F9 = 7.04 0.001 *** UVS phylogenetic MANOVA approx-F9 = 3.01 < 0.001 *** against a leaf PERMANOVA F9 = 7.07 0.001 *** VS phylogenetic MANOVA approx-F9 = 3.10 < 0.001 *** xyzL woodland shade PERMANOVA F9 = 5.33 0.001 *** UVS phylogenetic MANOVA approx-F9 = 3.12 < 0.001 *** against the sky PERMANOVA F9 = 5.34 0.002 ** VS phylogenetic MANOVA approx-F9 = 3.39 < 0.001 *** PERMANOVA F9 = 7.24 0.001 *** UVS phylogenetic MANOVA approx-F9 = 3.00 < 0.001 *** against a leaf PERMANOVA F9 = 7.24 0.001 *** VS phylogenetic MANOVA approx-F9 = 3.07 < 0.001 *** large gap PERMANOVA F9 = 5.37 0.001 *** UVS phylogenetic MANOVA approx-F9 = 3.14 < 0.001 *** against the sky PERMANOVA F9 = 5.37 0.001 *** VS phylogenetic MANOVA approx-F9 = 3.38 < 0.001 *** x, y and z are the mean coordinates in the tetrahedral colour space of transparent areas for each species and L is the mean luminance. -

A Rapid Biological Assessment of the Upper Palumeu River Watershed (Grensgebergte and Kasikasima) of Southeastern Suriname
Rapid Assessment Program A Rapid Biological Assessment of the Upper Palumeu River Watershed (Grensgebergte and Kasikasima) of Southeastern Suriname Editors: Leeanne E. Alonso and Trond H. Larsen 67 CONSERVATION INTERNATIONAL - SURINAME CONSERVATION INTERNATIONAL GLOBAL WILDLIFE CONSERVATION ANTON DE KOM UNIVERSITY OF SURINAME THE SURINAME FOREST SERVICE (LBB) NATURE CONSERVATION DIVISION (NB) FOUNDATION FOR FOREST MANAGEMENT AND PRODUCTION CONTROL (SBB) SURINAME CONSERVATION FOUNDATION THE HARBERS FAMILY FOUNDATION Rapid Assessment Program A Rapid Biological Assessment of the Upper Palumeu River Watershed RAP (Grensgebergte and Kasikasima) of Southeastern Suriname Bulletin of Biological Assessment 67 Editors: Leeanne E. Alonso and Trond H. Larsen CONSERVATION INTERNATIONAL - SURINAME CONSERVATION INTERNATIONAL GLOBAL WILDLIFE CONSERVATION ANTON DE KOM UNIVERSITY OF SURINAME THE SURINAME FOREST SERVICE (LBB) NATURE CONSERVATION DIVISION (NB) FOUNDATION FOR FOREST MANAGEMENT AND PRODUCTION CONTROL (SBB) SURINAME CONSERVATION FOUNDATION THE HARBERS FAMILY FOUNDATION The RAP Bulletin of Biological Assessment is published by: Conservation International 2011 Crystal Drive, Suite 500 Arlington, VA USA 22202 Tel : +1 703-341-2400 www.conservation.org Cover photos: The RAP team surveyed the Grensgebergte Mountains and Upper Palumeu Watershed, as well as the Middle Palumeu River and Kasikasima Mountains visible here. Freshwater resources originating here are vital for all of Suriname. (T. Larsen) Glass frogs (Hyalinobatrachium cf. taylori) lay their -

Elevation in Tropical Sky Islands As the Common Driver in Structuring Genes and Communities of Freshwater Organisms
Research Collection Journal Article Elevation in tropical sky islands as the common driver in structuring genes and communities of freshwater organisms Author(s): Gueuning, Morgan; Suchan, Tomasz; Rutschmann, Sereina; Gattolliat, Jean-Luc; Jamsari, Jamsari; Kamil, Al I.; Pitteloud, Camille; Buerki, Sven; Balke, Michael; Sartori, Michel; Alvarez, Nadir Publication Date: 2017 Permanent Link: https://doi.org/10.3929/ethz-b-000217957 Originally published in: Scientific Reports 7(1), http://doi.org/10.1038/s41598-017-16069-y Rights / License: Creative Commons Attribution 4.0 International This page was generated automatically upon download from the ETH Zurich Research Collection. For more information please consult the Terms of use. ETH Library www.nature.com/scientificreports OPEN Elevation in tropical sky islands as the common driver in structuring genes and communities of Received: 30 January 2017 Accepted: 7 November 2017 freshwater organisms Published: xx xx xxxx Morgan Gueuning1,2, Tomasz Suchan1,12, Sereina Rutschmann1,3, Jean-Luc Gattolliat1,4, Jamsari Jamsari5, Al Ihsan Kamil5, Camille Pitteloud1,6,7, Sven Buerki8,9, Michael Balke10, Michel Sartori 1,4 & Nadir Alvarez 1,11 Tropical mountains are usually characterized by a vertically-arranged sequence of ecological belts, which, in contrast to temperate habitats, have remained relatively stable in space across the Quaternary. Such long-lasting patterning of habitats makes them ideal to test the role of environmental pressure in driving ecological and evolutionary processes. Using Sumatran freshwater mayfy communities, we test whether elevation, rather than other spatial factors (i.e. volcanoes, watersheds) structures both species within communities and genes within species. Based on the analysis of 31 mayfy (Ephemeroptera) communities and restriction-site-associated-DNA sequencing in the four most ubiquitous species, we found elevation as the major spatial component structuring both species and genes in the landscape. -

Rev Iss Web Mec 13773 25-22 5765..5784
Molecular Ecology (2016) 25, 5765–5784 doi: 10.1111/mec.13773 Into the Andes: multiple independent colonizations drive montane diversity in the Neotropical clearwing butterflies Godyridina NICOLAS CHAZOT,*† KEITH R. WILLMOTT,‡ FABIEN L. CONDAMINE,§¶ DONNA LISA DE-SILVA,* ANDREV.L.FREITAS,**GERARDOLAMAS,†† HELENE MORLON,‡‡ CARLOS E. GIRALDO,§§ CHRIS D. JIGGINS,¶¶ MATHIEU JORON,*** JAMES MALLET,††† SANDRA URIBE‡‡‡ and MARIANNE ELIAS* *Institut de Systematique, Evolution, Biodiversite, ISYEB – UMR 7205 – CNRS MNHN UPMC EPHE, Museum national d’Histoire naturelle, Sorbonne Universites, 57 rue Cuvier CP50, F-75005 Paris, France, †Department of Biology, University of Lund, 223 62 Lund, Sweden, ‡McGuire Center for Lepidoptera and Biodiversity, Florida Museum of Natural History, University of Florida, Gainesville, FL 32611, USA, §CNRS, UMR 5554 Institut des Sciences de l’Evolution (Universite de Montpellier), Place Eugene Bataillon, 34095 Montpellier, France, ¶Department of Biological Sciences, University of Alberta, T6G 2E9 Edmonton, AB, Canada, **Departamento de Zoologia and Museu de Zoologia, Instituto de Biologia, Universidade Estadual de Campinas, Campinas, S~ao Paulo, Brazil, ††Museo de Historia Natural, Universidad Nacional de San Marcos, Lima, Peru, ‡‡IBENS, Ecole Normale Superieure, UMR 8197 CNRS, Paris, France, §§Grupo de Investigacion de Sanidad Vegetal, Universidad Catolica de Oriente, Rionegro, Antioquia, Colombia, ¶¶Department of Zoology, University of Cambridge, Cambridge, UK, ***Centre d’Ecologie Fonctionnelle et Evolutive, CEFE, UMR 5175 CNRS – EPHE – Universite de Montpellier – Universite Paul Valery Montpellier, 34293 Montpellier 5, France, †††Department of Organismic and Evolutionary Biology, Harvard University, Cambridge, MA 02138, USA, ‡‡‡Universidad Nacional de Colombia, sede Medellın, Medellın, Colombia Abstract Understanding why species richness peaks along the Andes is a fundamental question in the study of Neotropical biodiversity. -

Molecular Phylogenetics of the Neotropical Butterfly Subtribe Oleriina
Molecular Phylogenetics and Evolution 55 (2010) 1032–1041 Contents lists available at ScienceDirect Molecular Phylogenetics and Evolution journal homepage: www.elsevier.com/locate/ympev Molecular phylogenetics of the neotropical butterfly subtribe Oleriina (Nymphalidae: Danainae: Ithomiini) Donna Lisa de-Silva a,*, Julia J. Day a, Marianne Elias b,c, Keith Willmott d, Alaine Whinnett a, James Mallet a a Department of Genetics, Evolution and Environment, University College London, Wolfson House, 4 Stephenson Way, London NW1 2HE, UK b Imperial College London, Silwood Park, Buckhurst Road, Ascot, Berkshire SL5 7PY, UK c CNRS, UMR 7205, Muséum National d’Histoire Naturelle, 45 Rue Buffon, CP50, 75005 Paris, France d McGuire Center for Lepidoptera, Florida Museum of Natural History, University of Florida, P.O. Box 112710, Gainesville, FL 32611-2710, USA article info abstract Article history: The Oleriina is one of the most speciose subtribes of the neotropical nymphalid butterfly tribe Ithomiini. Received 9 September 2009 They are widely distributed across the Andes and Amazonian lowlands and like other ithomiines they are Revised 22 December 2009 involved in complex mimicry rings. This subtribe is of particular interest because it contains the most Accepted 9 January 2010 diverse ithomiine genus, Oleria, as well as two genera, Megoleria and Hyposcada, that feed on hostplants Available online 15 January 2010 not utilized elsewhere in the tribe. Here we present the first comprehensive species-level phylogeny for the Oleriina, representing 83% of recognised species in the group, and based on 6698 bp from eight mito- Keywords: chondrial (mt) and nuclear (nc) genes. Topologies are largely congruent for ncDNA and the concatenated Lepidoptera dataset and the genera Oleria, Hyposcada and Megoleria are recovered and well-supported, although Speciation Phylogeny strongly discordant genealogy between mtDNA and ncDNA suggest possible introgression among Hypos- Hybridization cada and Megoleria. -

Illustrations of New Species of Exotic Butterflies
^^3 ILLUSTRATIONS OP NEW SPECIES OP EXOTIC BUTTERFLIES WILLIAM C. HBWITSON. lit SELECTED CHIEFLY FEOM HIS OWN COLLECTION. VOL. V. JOHN VAN VOORST, LONDON. 1872—1876. LONDON : PRINTED BY WOODFALL AND KINDER, . MILFORD LANE, STRAND, W.O. PREFACE. It is with regret, not however unmixed with satisfaction, tliat I come to the close of a work which has been to me a twenty-five years' labour of love. With regret that age and failing health forbid me to commence another volume. With satisfaction when I remember the great kindness which I have experienced personally from aU lepidopterists during its progress, and the very favourable reception it has met with from aE, and especially from those whose position as naturalists gives value to their opinions. I have many times during the progress of the book expressed my grati- tude for the kindness and liberality which I have met with from Dr. Boisduval, not only in giving me free access to his collections, but in allowing me during his absence to select from them and bring home with me aU that I wished to figure, feeling, as I do, the difiiculty I should myself experience in being parted from any of my treasures. To the generous encouragement met with from Mr. Wilson Saunders, especially at its commencement, the work owes much of its success. I am fully aware of and regret many errors, but have endeavoured to atone for them as much as possible by myself pointing them out and correcting them. It has always been my study to make the work useful rather than attractive.