Intersectional Hybridization Among Trichostema Species
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Redalyc.Géneros De Lamiaceae De México, Diversidad Y Endemismo
Revista Mexicana de Biodiversidad ISSN: 1870-3453 [email protected] Universidad Nacional Autónoma de México México Martínez-Gordillo, Martha; Fragoso-Martínez, Itzi; García-Peña, María del Rosario; Montiel, Oscar Géneros de Lamiaceae de México, diversidad y endemismo Revista Mexicana de Biodiversidad, vol. 84, núm. 1, marzo, 2013, pp. 30-86 Universidad Nacional Autónoma de México Distrito Federal, México Disponible en: http://www.redalyc.org/articulo.oa?id=42526150034 Cómo citar el artículo Número completo Sistema de Información Científica Más información del artículo Red de Revistas Científicas de América Latina, el Caribe, España y Portugal Página de la revista en redalyc.org Proyecto académico sin fines de lucro, desarrollado bajo la iniciativa de acceso abierto Revista Mexicana de Biodiversidad 84: 30-86, 2013 DOI: 10.7550/rmb.30158 Géneros de Lamiaceae de México, diversidad y endemismo Genera of Lamiaceae from Mexico, diversity and endemism Martha Martínez-Gordillo1, Itzi Fragoso-Martínez1, María del Rosario García-Peña2 y Oscar Montiel1 1Herbario de la Facultad de Ciencias, Facultad de Ciencias, Universidad Nacional Autónoma de México. partado postal 70-399, 04510 México, D.F., México. 2Herbario Nacional de México, Instituto de Biología, Universidad Nacional Autónoma de México. Apartado postal 70-367, 04510 México, D.F., México. [email protected] Resumen. La familia Lamiaceae es muy diversa en México y se distribuye con preferencia en las zonas templadas, aunque es posible encontrar géneros como Hyptis y Asterohyptis, que habitan en zonas secas y calientes; es una de las familias más diversas en el país, de la cual no se tenían datos actualizados sobre su diversidad y endemismo. -
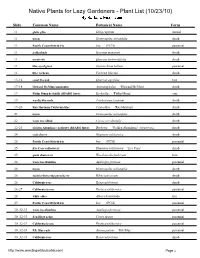
Native Plants for Lazy Gardeners - Plant List (10/23/10)
Native Plants for Lazy Gardeners - Plant List (10/23/10) Slide Common Name Botanical Name Form 11 globe gilia Gilia capitata annual 11 toyon Heteromeles arbutifolia shrub 11 Pacific Coast Hybrid iris Iris (PCH) perennial 11 goldenbush Isocoma menziesii shrub 11 scrub oak Quercus berberidifolia shrub 11 blue-eyed grass Sisyrinchium bellum perennial 11 lilac verbena Verbena lilacina shrub 13-16 coast live oak Quercus agrifolia tree 17-18 Howard McMinn man anita Arctostaphylos 'Howard McMinn' shrub 19 Philip Mun keckiella (RSABG Intro) Keckiella 'Philip Munz' ine 19 woolly bluecurls Trichostema lanatum shrub 19-20 Ray Hartman California lilac Ceanothus 'Ray Hartman' shrub 21 toyon Heteromeles arbutifolia shrub 22 western redbud Cercis occidentalis shrub 22-23 Golden Abundance barberry (RSABG Intro) Berberis 'Golden Abundance' (MAHONIA) shrub 2, coffeeberry Rhamnus californica shrub 25 Pacific Coast Hybrid iris Iris (PCH) perennial 25 Eve Case coffeeberry Rhamnus californica '. e Case' shrub 25 giant chain fern Woodwardia fimbriata fern 26 western columbine Aquilegia formosa perennial 26 toyon Heteromeles arbutifolia shrub 26 fuchsia-flowering gooseberry Ribes speciosum shrub 26 California rose Rosa californica shrub 26-27 California fescue Festuca californica perennial 28 white alder Alnus rhombifolia tree 29 Pacific Coast Hybrid iris Iris (PCH) perennial 30 032-33 western columbine Aquilegia formosa perennial 30 032-33 San Diego sedge Carex spissa perennial 30 032-33 California fescue Festuca californica perennial 30 032-33 Elk Blue rush Juncus patens '.l1 2lue' perennial 30 032-33 California rose Rosa californica shrub http://www weedingwildsuburbia com/ Page 1 30 032-3, toyon Heteromeles arbutifolia shrub 30 032-3, fuchsia-flowering gooseberry Ribes speciosum shrub 30 032-3, Claremont pink-flowering currant (RSA Intro) Ribes sanguineum ar. -

Well-Known Plants in Each Angiosperm Order
Well-known plants in each angiosperm order This list is generally from least evolved (most ancient) to most evolved (most modern). (I’m not sure if this applies for Eudicots; I’m listing them in the same order as APG II.) The first few plants are mostly primitive pond and aquarium plants. Next is Illicium (anise tree) from Austrobaileyales, then the magnoliids (Canellales thru Piperales), then monocots (Acorales through Zingiberales), and finally eudicots (Buxales through Dipsacales). The plants before the eudicots in this list are considered basal angiosperms. This list focuses only on angiosperms and does not look at earlier plants such as mosses, ferns, and conifers. Basal angiosperms – mostly aquatic plants Unplaced in order, placed in Amborellaceae family • Amborella trichopoda – one of the most ancient flowering plants Unplaced in order, placed in Nymphaeaceae family • Water lily • Cabomba (fanwort) • Brasenia (watershield) Ceratophyllales • Hornwort Austrobaileyales • Illicium (anise tree, star anise) Basal angiosperms - magnoliids Canellales • Drimys (winter's bark) • Tasmanian pepper Laurales • Bay laurel • Cinnamon • Avocado • Sassafras • Camphor tree • Calycanthus (sweetshrub, spicebush) • Lindera (spicebush, Benjamin bush) Magnoliales • Custard-apple • Pawpaw • guanábana (soursop) • Sugar-apple or sweetsop • Cherimoya • Magnolia • Tuliptree • Michelia • Nutmeg • Clove Piperales • Black pepper • Kava • Lizard’s tail • Aristolochia (birthwort, pipevine, Dutchman's pipe) • Asarum (wild ginger) Basal angiosperms - monocots Acorales -

Outline of Angiosperm Phylogeny
Outline of angiosperm phylogeny: orders, families, and representative genera with emphasis on Oregon native plants Priscilla Spears December 2013 The following listing gives an introduction to the phylogenetic classification of the flowering plants that has emerged in recent decades, and which is based on nucleic acid sequences as well as morphological and developmental data. This listing emphasizes temperate families of the Northern Hemisphere and is meant as an overview with examples of Oregon native plants. It includes many exotic genera that are grown in Oregon as ornamentals plus other plants of interest worldwide. The genera that are Oregon natives are printed in a blue font. Genera that are exotics are shown in black, however genera in blue may also contain non-native species. Names separated by a slash are alternatives or else the nomenclature is in flux. When several genera have the same common name, the names are separated by commas. The order of the family names is from the linear listing of families in the APG III report. For further information, see the references on the last page. Basal Angiosperms (ANITA grade) Amborellales Amborellaceae, sole family, the earliest branch of flowering plants, a shrub native to New Caledonia – Amborella Nymphaeales Hydatellaceae – aquatics from Australasia, previously classified as a grass Cabombaceae (water shield – Brasenia, fanwort – Cabomba) Nymphaeaceae (water lilies – Nymphaea; pond lilies – Nuphar) Austrobaileyales Schisandraceae (wild sarsaparilla, star vine – Schisandra; Japanese -

Vascular Plant and Vertebrate Inventory of Chiricahua National Monument
In Cooperation with the University of Arizona, School of Natural Resources Vascular Plant and Vertebrate Inventory of Chiricahua National Monument Open-File Report 2008-1023 U.S. Department of the Interior U.S. Geological Survey National Park Service This page left intentionally blank. In cooperation with the University of Arizona, School of Natural Resources Vascular Plant and Vertebrate Inventory of Chiricahua National Monument By Brian F. Powell, Cecilia A. Schmidt, William L. Halvorson, and Pamela Anning Open-File Report 2008-1023 U.S. Geological Survey Southwest Biological Science Center Sonoran Desert Research Station University of Arizona U.S. Department of the Interior School of Natural Resources U.S. Geological Survey 125 Biological Sciences East National Park Service Tucson, Arizona 85721 U.S. Department of the Interior DIRK KEMPTHORNE, Secretary U.S. Geological Survey Mark Myers, Director U.S. Geological Survey, Reston, Virginia: 2008 For product and ordering information: World Wide Web: http://www.usgs.gov/pubprod Telephone: 1-888-ASK-USGS For more information on the USGS-the Federal source for science about the Earth, its natural and living resources, natural hazards, and the environment: World Wide Web:http://www.usgs.gov Telephone: 1-888-ASK-USGS Suggested Citation Powell, B.F., Schmidt, C.A., Halvorson, W.L., and Anning, Pamela, 2008, Vascular plant and vertebrate inventory of Chiricahua National Monument: U.S. Geological Survey Open-File Report 2008-1023, 104 p. [http://pubs.usgs.gov/of/2008/1023/]. Cover photo: Chiricahua National Monument. Photograph by National Park Service. Note: This report supersedes Schmidt et al. (2005). Any use of trade, product, or firm names is for descriptive purposes only and does not imply endorsement by the U.S. -

Vascular Plant and Vertebrate Inventory of Fort Bowie National Historic Site Vascular Plant and Vertebrate Inventory of Fort Bowie National Historic Site
Powell, Schmidt, Halvorson In Cooperation with the University of Arizona, School of Natural Resources Vascular Plant and Vertebrate Inventory of Fort Bowie National Historic Site Vascular Plant and Vertebrate Inventory of Fort Bowie National Historic Site Plant and Vertebrate Vascular U.S. Geological Survey Southwest Biological Science Center 2255 N. Gemini Drive Flagstaff, AZ 86001 Open-File Report 2005-1167 Southwest Biological Science Center Open-File Report 2005-1167 February 2007 U.S. Department of the Interior U.S. Geological Survey National Park Service In cooperation with the University of Arizona, School of Natural Resources Vascular Plant and Vertebrate Inventory of Fort Bowie National Historic Site By Brian F. Powell, Cecilia A. Schmidt , and William L. Halvorson Open-File Report 2005-1167 December 2006 USGS Southwest Biological Science Center Sonoran Desert Research Station University of Arizona U.S. Department of the Interior School of Natural Resources U.S. Geological Survey 125 Biological Sciences East National Park Service Tucson, Arizona 85721 U.S. Department of the Interior DIRK KEMPTHORNE, Secretary U.S. Geological Survey Mark Myers, Director U.S. Geological Survey, Reston, Virginia: 2006 For product and ordering information: World Wide Web: http://www.usgs.gov/pubprod Telephone: 1-888-ASK-USGS For more information on the USGS-the Federal source for science about the Earth, its natural and living resources, natural hazards, and the environment: World Wide Web:http://www.usgs.gov Telephone: 1-888-ASK-USGS Suggested Citation Powell, B. F, C. A. Schmidt, and W. L. Halvorson. 2006. Vascular Plant and Vertebrate Inventory of Fort Bowie National Historic Site. -

Plant Breading
SNA Research Conference Vol. 52 2007 Plant Breeding and Evaluation Tom Ranney Section Editor and Moderator Plant Breeding and Evaluation Section 326 SNA Research Conference Vol. 52 2007 New Callicarpa Species with Breeding Potential Ryan N. Contreras and John M. Ruter University of Georgia, Dept. of Horticulture, Tifton, GA 31793 [email protected] Index Words: beautyberry, species evaluation, ornamental plant breeding Significance to Industry: There is a great deal of available Callicarpa L. germplasm that has yet to be utilized by the nursery industry in the U.S. Taxa currently being evaluated are likely to have potential as breeding material or direct commercial marketability. With new breeding material and selections for introduction the number of beautyberry cultivars for use in southeastern gardens has the potential to expand greatly. Nature of Work: Callicarpa L. is a genus of ~150 species of shrubs and trees distributed throughout the world including warm-temperate and tropical America, SE Asia, Malaysia, Pacific Islands, and Australia (5) with the greatest concentration of species found in SE Asia, specifically the Philippine Islands (1). Of the New World species the highest concentration occurs in Cuba, with ~20 native species (1). There are currently four species commonly found in cultivation in the U.S.: C. americana L., C. bodinieri Lév., C. dichotoma (Lour.)K.Koch, and C. japonica Thunb. with a limited number of varieties or cultivars of each to choose from (3). Beautyberries, desired primarily for their handsome berries produced in fall, have been selected for white-fruiting varieties, finer textured varieties, increased berry production, and variegated foliage. -

Biological Resources Assessment the Ranch ±530- Acre Study Area City of Rancho Cordova, California
Biological Resources Assessment The Ranch ±530- Acre Study Area City of Rancho Cordova, California Prepared for: K. Hovnanian Homes October 13, 2017 Prepared by: © 2017 TABLE OF CONTENTS 1.0 Introduction ......................................................................................................................... 1 1.1. Project Description ........................................................................................................... 1 2.0 Regulatory Framework ........................................................................................................ 2 2.1. Federal Regulations .......................................................................................................... 2 2.1.1. Federal Endangered Species Act ............................................................................... 2 2.1.2. Migratory Bird Treaty Act ......................................................................................... 2 2.1.3. The Bald and Golden Eagle Protection Act ............................................................... 2 2.2. State Jurisdiction .............................................................................................................. 3 2.2.1. California Endangered Species Act ........................................................................... 3 2.2.2. California Department of Fish and Game Codes ...................................................... 3 2.2.3. Native Plant Protection Act ..................................................................................... -

Draft Plant Propagation Protocol
Plant Propagation Protocol for Trichostema oblongum Benth. ESRM 412 – Native Plant Production Protocol URL: https://courses.washington.edu/esrm412/protocols/TROB.pdf [3] [9] [5] TAXONOMY Plant Family Scientific Name Lamiaceae [1] Common Name Mint family [2] Species Scientific Name Scientific Name Trichostema oblongum Benth. [1] Varieties None [1, 3, 4] Sub-species None [1, 3, 4] Cultivar None [1, 3, 4] Common None [1, 3, 4] Synonym(s) Common Oblong bluecurls, Mountain bluecurls [1, 3, 4, 5] Name(s) Species Code (as TROB [1] per USDA Plants database) GENERAL INFORMATION Geographical Confined to Western North America [1, 3, 4, 6]. Found as far south as Kern range County, CA [1, 5, 7]; and as far north as Castlegar, B.C [6, 8]. In WA, only found east of the Cascades [1, 3, 9]. [1] Ecological Commonly found along dry margins of vernally moist meadows, distribution streambanks, and forest openings, often on disturbed and/or alkaline soils [3, 4, 5, 6, 9] Climate and Grows in vernally moist arid regions, at elevations of 100–3,000 meters elevation range (330–9,840 ft); confined to 600-730 meters (1,950-2,400 ft) in Washington [3, 4, 5, 6, 9] Local habitat Somewhat common in northern CA, the Willamette Valley of OR, and along and abundance the eastern slopes of the Sierra Nevada [1, 4, 5, 7]. Rare in WA, ID, & B.C. [1, 3, 6, 8, 9] In CA and OR, often associated with forest clearings in Yellow pine forests, Red Fir Forests, Lodgepole pine Forests, Subalpine Forests, California mixed evergreen forests, and North Coastal Coniferous Forests [5, 7] In WA, often associated with Navarettia intertexta, Deschampsia danthonioides, Agrostis interrupta, Madia minima, Juncus bufonius, Juncus tiehmii, Orthocarpus tenuifolius, Trifolium variegatum, Mimulus brevifolius, Epilobium minutum, and Artemesia tridentate [9]. -

Bidens Laevis Bur's Marigold
Gillett Plant Palette Back to Natives Restoration, a 501(c)3 backtonatives.org Back to Natives Mission: To encourage and actively participate in the conservation and restoration of Orange County and California wildlands, through education and habitat restoration programs featuring native plants and biodiversity as a centralizing theme. Biodiversity or biological diversity is the variation of life forms within a given ecosystem, biome or for the entire Earth. Biodiversity measures the health of biological systems (historical species present vs. historical species extinct). What does non-native mean? When non-native plants spread and become established in wild areas they out-compete native plants. Diseases or weather conditions which kept the plants in check in their homeland are absent here. These weeds deprive our wild animals of food and shelter, since native animals cannot make much use of them. What does native mean? “Natives” were here prior to European contact. California's native plants & animals evolved here over a very long period, and form a complex network of relationships. They are adapted to our climate and geography. Why Grow Native Plants? Promote Biodiversity • In California, there are over 6,000 native plant species, more than in the central and northeastern US and Canada combined. • More than 1500 of these plant species are endemic to (found only in) California, and most of these endemic species are found in Southern California. • Southern California is one of the 25 global biodiversity “hotspots.” Hotspots are where the largest number of different species can be found, especially those species found nowhere else. • More than 60 percent of the Earth's total species live in hotspots, which cover only 1.44 percent of its surface. -

Griffith Park Rare Plant Survey
Cooper Ecological Monitoring, Inc. EIN 72-1598095 Daniel S. Cooper, President 5850 W. 3rd St. #167 Los Angeles, CA 90036 (323) 397-3562 [email protected] Griffith Park Rare Plant Survey Plummer's mariposa-lily Calochortus plummerae (CNPS 1B.2) blooms near Skyline Trail in the northeastern corner of Griffith Park, 26 May 2010 (ph. DSC). Prepared by: Daniel S. Cooper Cooper Ecological Monitoring, Inc. October 2010 1 Part I. Summary of Findings Part II (species accounts) begins after p. 26. We present information on extant occurrences of 15 special-status species, subspecies and/or varieties of vascular plants in Griffith Park and contiguous open space, including three for which no known local specimen existed prior to this study: slender mariposa-lily (Calochortus clavatus var. gracilis; CNPS 1B.2), Humboldt lily (Lilium humboldtii var. ocellatum; CNPS 4.2), and Hubby's phacelia (Phacelia hubbyi; CNPS 4.2). Using lists developed by local botanists, we document - from specimens or digital photographs - extant occurrences of nearly 40 additional plant taxa felt to be of conservation concern in the eastern Santa Monica Mountains, including 16 for which no prior specimen existed for the park or surrounding open space. We also identify several dozen taxa known from the specimen record but unconfirmed in the park in recent years. From this information, we discuss patterns of occurrence of rare plants in the park, drawing attention to "hotspots" for rare species diversity, such as Spring Canyon and Royce Canyon, and identify areas, particularly in the northeastern corner of the park and along the southeastern border, where rare plants are relatively poorly represented in the landscape. -

Native Plants for Hummingbirds
NATIVE PLANTS FOR HUMMINGBIRDS Theodore Payne Foundation for Wild Flowers and Native Plants 10459 Tuxford Street, Sun Valley, CA 91352, (818) 768-1802, theodorepayne.org Hummingbirds are attracted to flowers in the red spectrum but are also attracted to many other colors, including white, purple and yellow. They prefer red flowers because many insects avoid red blossoms, thus there is more nectar in them. The typical hummingbird flower is a long flared tube that keeps out rival pollinators and invites the little birds to lap up the sweet nectar with their long bills and tongues. To provide food year round, plant an assortment of native plants with different blooming seasons. Avoid using pesticides, as hummingbirds subsist primarily on small insects during fall and winter, and hummers use spider webs to build their nests. Some garden plants provide nesting material, such as the fuzzy hairs on the underside of sycamore leaves. A water source is appreciated; shallow, moving water is best. Several species of hummingbirds are year-round residents in Southern California; others migrate through our region. These nectar plants will attract and support hummingbirds throughout the seasons. SPRING Aquilegia formosa (western columbine) – 2-3’ H x 1-2’ W – Showy red and yellow flowers. Sun or shade. Berberis (Mahonia) species and cultivars (barberry, Oregon grape) – Sizes vary – Barrier plants with prickly foliage and yellow flowers. Calliandra californica (red fairyduster) – 5’ H x 5’ W – Showy Baja CA native; blooms year round. Cirsium occidentale (California thistle) – 4’ H x 1’ W – Annual with magenta flowers that attract many pollinators. Cobweb-like flower filaments are used for nesting.