Silurian Vertebrate Remains from the Oslo Region, Norway, and Their Implications for Regional Biostratigraphy
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Bioforsk-Rapport-2009-04-71.Pdf (1.987Mb)
”Mat fra Toten” Bioforsk Rapport Bioforsk Report Vol. 4 Nr. 71 2009 Jordsmonn, klima og andre forutsetninger for kvalitetsproduksjon av mat på Toten Hans Stabbetorp Bioforsk Øst Apelsvoll Foto: Unni Abrahamsen Hovedkontor/Head office Bioforsk Øst Apelsvoll Frederik A. Dahls vei 20 Bioforsk Arable Crops Division N-1432 Ås 2849 Kapp Tel.: (+47) 40 60 41 00 Tel.: (+47) 40 60 41 00 [email protected] [email protected] Tittel/Title: Jordsmonn, klima og andre forutsetninger for kvalitetsproduksjon av mat på Toten Forfatter(e)/Author(s): Hans Stabbetorp Dato/Date: Tilgjengelighet/Availability: Prosjekt nr./Project No.: Saksnr./Archive No.: 08.06.2009 Åpen 1210155 621.5 Rapport nr./Report No.: ISBN-nr./ISBN-no: Antall sider/Number of Antall vedlegg/Number pages: of appendices: 4(71) 2009 978–82–17–00508-7 31 2 Oppdragsgiver/Employer: Kontaktperson/Contact person: ”Mat fra Toten” Per Odd Gjestvang og Ole Anton Hoel Stikkord/Keywords: Fagområde/Field of work: Toten, jordsmonn, klima, kvalitetsproduksjon av mat Korn, potet og grønnsaker Sammendrag: Publikasjonen omhandler geologi og jordsmonnsutvikling, klima og andre forutsetninger for kvalitetsproduksjon av mat på Toten. Se for øvrig omfattende sammendrag side 29-30. Land/Country: Norge Fylke/County: Oppland Kommune/Municipality: Østre Toten Godkjent / Approved Prosjektleder / Project leader Ingvar Hage Hans Stabbetorp Innhold Toten………………………………………………………………………………………………………………………… 3 Østre Toten……………………………………………………………………………………………………… 3 Vestre Toten…………………………………………………………………………………………………… 3 Jordsmonnet -

Early Silurian Oceanic Episodes and Events
Journal of the Geological Society, London, Vol. 150, 1993, pp. 501-513, 3 figs. Printed in Northern Ireland Early Silurian oceanic episodes and events R. J. ALDRIDGE l, L. JEPPSSON 2 & K. J. DORNING 3 1Department of Geology, The University, Leicester LE1 7RH, UK 2Department of Historical Geology and Palaeontology, SiSlvegatan 13, S-223 62 Lund, Sweden 3pallab Research, 58 Robertson Road, Sheffield $6 5DX, UK Abstract: Biotic cycles in the early Silurian correlate broadly with postulated sea-level changes, but are better explained by a model that involves episodic changes in oceanic state. Primo episodes were characterized by cool high-latitude climates, cold oceanic bottom waters, and high nutrient supply which supported abundant and diverse planktonic communities. Secundo episodes were characterized by warmer high-latitude climates, salinity-dense oceanic bottom waters, low diversity planktonic communities, and carbonate formation in shallow waters. Extinction events occurred between primo and secundo episodes, with stepwise extinctions of taxa reflecting fluctuating conditions during the transition period. The pattern of turnover shown by conodont faunas, together with sedimentological information and data from other fossil groups, permit the identification of two cycles in the Llandovery to earliest Weniock interval. The episodes and events within these cycles are named: the Spirodden Secundo episode, the Jong Primo episode, the Sandvika event, the Malm#ykalven Secundo episode, the Snipklint Primo episode, and the lreviken event. Oceanic and climatic cyclicity is being increasingly semblages (Johnson et al. 1991b, p. 145). Using this recognized in the geological record, and linked to major and approach, they were able to detect four cycles within the minor sedimentological and biotic fluctuations. -

Categorical Versus Geometric Morphometric Approaches To
[Palaeontology, 2020, pp. 1–16] CATEGORICAL VERSUS GEOMETRIC MORPHOMETRIC APPROACHES TO CHARACTERIZING THE EVOLUTION OF MORPHOLOGICAL DISPARITY IN OSTEOSTRACI (VERTEBRATA, STEM GNATHOSTOMATA) by HUMBERTO G. FERRON 1,2* , JENNY M. GREENWOOD1, BRADLEY DELINE3,CARLOSMARTINEZ-PEREZ 1,2,HECTOR BOTELLA2, ROBERT S. SANSOM4,MARCELLORUTA5 and PHILIP C. J. DONOGHUE1,* 1School of Earth Sciences, University of Bristol, Life Sciences Building, Tyndall Avenue, Bristol, BS8 1TQ, UK; [email protected], [email protected], [email protected] 2Institut Cavanilles de Biodiversitat i Biologia Evolutiva, Universitat de Valencia, C/ Catedratic Jose Beltran Martınez 2, 46980, Paterna, Valencia, Spain; [email protected], [email protected] 3Department of Geosciences, University of West Georgia, Carrollton, GA 30118, USA; [email protected] 4School of Earth & Environmental Sciences, University of Manchester, Manchester, M13 9PT, UK; [email protected] 5School of Life Sciences, University of Lincoln, Riseholme Hall, Lincoln, LN2 2LG, UK; [email protected] *Corresponding authors Typescript received 2 October 2019; accepted in revised form 27 February 2020 Abstract: Morphological variation (disparity) is almost aspects of morphology. Phylomorphospaces reveal conver- invariably characterized by two non-mutually exclusive gence towards a generalized ‘horseshoe’-shaped cranial mor- approaches: (1) quantitatively, through geometric morpho- phology and two strong trends involving major groups of metrics; -

Oslo Government District Bombing and Utøya Island Shooting July 22, 2011: the Immediate Prehospital Emergency Medical Service Response Sollid Et Al
Oslo government district bombing and Utøya island shooting July 22, 2011: The immediate prehospital emergency medical service response Sollid et al. Sollid et al. Scandinavian Journal of Trauma, Resuscitation and Emergency Medicine 2012, 20:3 http://www.sjtrem.com/content/20/1/3 (17 January 2012) Sollid et al. Scandinavian Journal of Trauma, Resuscitation and Emergency Medicine 2012, 20:3 http://www.sjtrem.com/content/20/1/3 ORIGINALRESEARCH Open Access Oslo government district bombing and Utøya island shooting July 22, 2011: The immediate prehospital emergency medical service response Stephen JM Sollid1,2,3*, Rune Rimstad4,5,6, Marius Rehn2, Anders R Nakstad1, Ann-Elin Tomlinson7, Terje Strand1, Hans Julius Heimdal1, Jan Erik Nilsen1,8 and Mårten Sandberg1,9, for Collaborating group Abstract Background: On July 22, 2011, a single perpetrator killed 77 people in a car bomb attack and a shooting spree incident in Norway. This article describes the emergency medical service (EMS) response elicited by the two incidents. Methods: A retrospective and observational study was conducted based on data from the EMS systems involved and the public domain. The study was approved by the Data Protection Official and was defined as a quality improvement project. Results: We describe the timeline and logistics of the EMS response, focusing on alarm, dispatch, initial response, triage and evacuation. The scenes in the Oslo government district and at Utøya island are described separately. Conclusions: Many EMS units were activated and effectively used despite the occurrence of two geographically separate incidents within a short time frame. Important lessons were learned regarding triage and evacuation, patient flow and communication, the use of and need for emergency equipment and the coordination of helicopter EMS. -
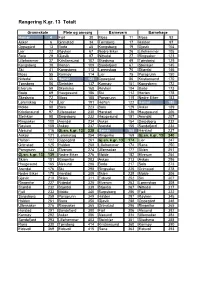
Rangering K.Gr. 13 Totalt
Rangering K.gr. 13 Totalt Grunnskole Pleie og omsorg Barnevern Barnehage Hamar 4 Fjell 30 Moss 11 Moss 92 Asker 6 Grimstad 34 Tønsberg 17 Halden 97 Oppegård 13 Bodø 45 Kongsberg 19 Gjøvik 104 Lier 22 Røyken 67 Nedre Eiker 26 Lillehammer 105 Sola 29 Gjøvik 97 Nittedal 27 Ringsaker 123 Lillehammer 37 Kristiansund 107 Skedsmo 49 Tønsberg 129 Kongsberg 38 Horten 109 Sandefjord 67 Steinkjer 145 Ski 41 Kongsberg 113 Lørenskog 70 Stjørdal 146 Moss 55 Karmøy 114 Lier 75 Porsgrunn 150 Nittedal 55 Hamar 123 Oppegård 86 Kristiansund 170 Tønsberg 56 Steinkjer 137 Karmøy 101 Kongsberg 172 Elverum 59 Skedsmo 168 Røyken 104 Bodø 173 Bodø 69 Haugesund 186 Ski 112 Horten 178 Skedsmo 72 Moss 188 Porsgrunn 115 Nedre Eiker 183 Lørenskog 74 Lier 191 Horten 122 Hamar 185 Molde 88 Sola 223 Sola 129 Asker 189 Kristiansund 97 Ullensaker 230 Harstad 136 Haugesund 206 Steinkjer 98 Sarpsborg 232 Haugesund 151 Arendal 207 Ringsaker 100 Arendal 234 Asker 154 Sarpsborg 232 Røyken 108 Askøy 237 Arendal 155 Sandefjord 234 Ålesund 116 Gj.sn. k.gr. 13 238 Hamar 168 Harstad 237 Askøy 121 Lørenskog 254 Ringerike 169 Gj.sn. k.gr. 13 240 Horten 122 Oppegård 261 Gj.sn. k.gr. 13 174 Lier 247 Grimstad 125 Halden 268 Lillehammer 174 Rana 250 Porsgrunn 133 Elverum 274 Ullensaker 177 Skien 251 Gj.sn. k.gr. 13 139 Nedre Eiker 276 Molde 182 Elverum 254 Skien 151 Ringerike 283 Askøy 213 Askøy 256 Haugesund 165 Ålesund 288 Bodø 217 Sola 273 Arendal 176 Ski 298 Ringsaker 225 Grimstad 278 Nedre Eiker 179 Harstad 309 Skien 239 Molde 306 Gjøvik 210 Skien 311 Eidsvoll 252 Ski 307 Ringerike -

200 Buss Rutetabell & Linjerutekart
200 buss rutetabell & linjekart 200 Hønefoss Vis I Nettsidemodus 200 buss Linjen Hønefoss har 5 ruter. For vanlige ukedager, er operasjonstidene deres 1 Hønefoss 06:00 - 23:40 2 Hønefoss Ekspress 14:55 - 16:25 3 Oslo 04:30 - 22:00 4 Oslo Ekspress 05:55 - 07:25 5 Sundvollen E16 14:58 Bruk Moovitappen for å ƒnne nærmeste 200 buss stasjon i nærheten av deg og ƒnn ut når neste 200 buss ankommer. Retning: Hønefoss 200 buss Rutetabell 38 stopp Hønefoss Rutetidtabell VIS LINJERUTETABELL mandag 06:00 - 23:40 tirsdag 06:00 - 23:40 Oslo Bussterminal 19, Oslo onsdag 06:00 - 23:40 St. Olavs Plass torsdag 06:00 - 23:40 Pilestredet, Oslo fredag 06:00 - 23:40 Vika Atrium lørdag 08:40 - 23:40 Munkedamsveien 53B, Oslo søndag 09:40 - 23:40 Filipstadveien Sjølyst Sjølystveien, Oslo 200 buss Info Vækerø Retning: Hønefoss Stopp: 38 Lysaker Stasjon Reisevarighet: 82 min Drammensveien 305, Oslo Linjeoppsummering: Oslo Bussterminal, St. Olavs Plass, Vika Atrium, Filipstadveien, Sjølyst, Vækerø, Oksenøyveien Lysaker Stasjon, Oksenøyveien, Ramstadsletta, Oksenøystien 2, Oslo Sandvika Bussterminal, Rustan, Brenna, Skoglund, Avtjerna, Solbakken, Sollihøgda, Sollihøgda Nedre, Ramstadsletta Homledal, Rørvik, Tyrifjord, Sundvollen E16, Drammensveien 503, Oslo Kroksund, Paddevika, Vik E16, Steinsåsen E16, Høyenhall E16, Steinsalléen, Vollgata, Sonerud, Gihle, Sandvika Bussterminal Botilrud, Hønensvingen E16, Tandberg, Trøgstad, Folangerveien 42, Oslo Ringerike Sykehus E16, Dalsbråten, Eikli, Hønefoss Sentrum Rustan Brenna Skoglund Avtjerna Ringeriksveien 459, Norway -

Guide to South Dakota Norwegian-American Collections
GUIDE TO COLLECTIONS RELATING TO SOUTH DAKOTA NORWEGIAN-AMERICANS Compiled by Harry F. Thompson, Ph.D. Director of Research Collections and Publications The Center for Western Studies With the assistance of Arthur R. Huseboe, Ph.D. and Paul B. Olson Additional assistance by Carol Riswold, D. Joy Harris, and Laura Plowman Originally published in 1991 by The Center for Western Studies, Augustana College, Sioux Falls, SD 57197 and updated in 2007. Original publication was made possible by a grant from the South Dakota Committee on the Humanities and by a gift from Harold L. Torness of Sisseton, South Dakota. TABLE OF CONTENTS Introduction 1 Albright College 2 Augustana College, The Center for Western Studies 3 Augustana College, Mikkelsen Library 4 Augustana College (IL), Swenson Swedish Immigration Research Center 5 Black Hills State University 6 Brookings Public Library 7 Canton Public Library 8 Centerville Public Library 9 Codington County Historical Society 10 Cornell University Libraries 11 Dakota State University 12 Dakota Wesleyan University 13 Dewey County Library 14 Elk Point Community Library 15 Grant County Public Library 16 Phoebe Apperson Hearst Library 17 J. Roland Hove 18 Luther College 19 Minnehaha County Historical Society 20 Minnehaha County Rural Public Library 21 Minnesota Historical Society, Research Center 2 22 Mitchell Area Genealogical Society 23 Mobridge Public Library 24 National Archives--Central Plains Region 25 North Dakota State University, North Dakota Institute for Regional Studies 26 Norwegian American Historical Association 27 James B. Olson 28 Rapid City Public Library 29 Rapid City Sons of Norway Borgund Lodge I-532 30 Regional Center for Mission--Region III, ELCA 31 St. -

Proposed Black Country UNESCO Global Geopark
Great things to see and do in the Proposed Black Country UNESCO Global Geopark Black Country UNESCO Global Geopark Project The layers lying above these are grey muddy Welcome to the world-class rocks that contain seams of ironstone, fireclay heritage which is the Black and coal with lots of fossils of plants and insects. These rocks tell us of a time some 310 million Country years ago (called the Carboniferous Period, The Black Country is an amazing place with a named after the carbon in the coal) when the captivating history spanning hundreds of Black Country was covered in huge steamy millions of years. This is a geological and cultural rainforests. undiscovered treasure of the UK, located at the Sitting on top of those we find reddish sandy heart of the country. It is just 30 minutes from rocks containing ancient sand dunes and Birmingham International Airport and 10 minutes pebbly river beds. This tells us that the landscape by train from the city of Birmingham. dried out to become a scorching desolate The Black Country is where many essential desert (this happened about 250 million years aspects of the Industrial Revolution began. It ago and lasted through the Permian and Triassic was the world’s first large scale industrial time periods). landscape where anything could be made, The final chapter in the making of our landscape earning it the nick-name the ‘workshop of the is often called the’ Ice Age’. It spans the last 2.6 world’ during the Industrial Revolution. This million years of our history when vast ice sheets short guidebook introduces some of the sites scraped across the surface of the area, leaving and features that are great things to see and a landscaped sculpted by ice and carved into places to explore across many parts of The the hills and valleys we see today. -

Fish and Amphibians
Fish and Amphibians Geology 331 Paleontology Phylum Chordata: Subphyla Urochordata, Cephalochordata, and: Subphylum Vertebrata Class Agnatha: jawless fish, includes the hagfish, conodonts, lampreys, and ostracoderms (armored jawless fish) Gnathostomates: jawed fish Class Chondrichthyes: cartilaginous fish Class Placoderms: armored fish Class Osteichthyes: bony fish Subclass Actinopterygians: ray-finned fish Subclass Sarcopterygians: lobe-finned fish Order Dipnoans: lung fish Order Crossopterygians: coelocanths and rhipidistians Class Amphibia Urochordates: Sea Squirts. Adults have a pharynx with gill slits. Larval forms are free-swimming and have a notochord. Chordates are thought to have evolved from the larval form by precocious sexual maturation. Chordate evolution Cephalochordate: Branchiostoma, the lancelet Pikaia, a cephalochordate from the Burgess Shale Yunnanozoon, a cephalochordate from the Lower Cambrian of China Haikouichthys, agnathan, Lower Cambrian of China - Chengjiang fauna, scale is 5 mm A living jawless fish, the lamprey, Class Agnatha Jawless fish do have teeth! A fossil jawless fish, Class Agnatha, Ostracoderm, Hemicyclaspis, Silurian Agnathan, Ostracoderm, Athenaegis, Silurian of Canada Agnathan, Ostracoderm, Pteraspis, Devonian of the U.K. Agnathan, Ostracoderm, Liliaspis, Devonian of Russia Jaws evolved by modification of the gill arch bones. The placoderms were the armored fish of the Paleozoic Placoderm, Dunkleosteus, Devonian of Ohio Asterolepis, Placoderms, Devonian of Latvia Placoderm, Devonian of Australia Chondrichthyes: A freshwater shark of the Carboniferous Fossil tooth of a Great White shark Chondrichthyes, Great White Shark Chondrichthyes, Carcharhinus Sphyrna - hammerhead shark Himantura - a ray Manta Ray Fish Anatomy: Ray-finned fish Osteichthyes: ray-finned fish: clownfish Osteichthyes: ray-finned fish, deep water species Lophius, an Eocene fish showing the ray fins. This is an anglerfish. -

Silurian Thelodonts from the Niur Formation, Central Iran
Silurian thelodonts from the Niur Formation, central Iran VACHIK HAIRAPETIAN, HENNING BLOM, and C. GILES MILLER Hairapetian, V., Blom, H., and Miller, C.G. 2008. Silurian thelodonts from the Niur Formation, central Iran. Acta Palaeontologica Polonica 53 (1): 85–95. Thelodont scales are described from the Silurian Niur Formation in the Derenjal Mountains, east central Iran. The mate− rial studied herein comes from four stratigraphic levels, composed of rocks formed in a shallow water, carbonate ramp en− vironment. The fauna includes a new phlebolepidiform, Niurolepis susanae gen. et sp. nov. of late Wenlock/?early Lud− low age and a late Ludlow loganelliiform, Loganellia sp. cf. L. grossi, which constitute the first record of these thelodont groups from Gondwana. The phlebolepidiform Niurolepis susanae gen. et sp. nov. is diagnosed by having trident trunk scales with a raised medial crown area separated by two narrow spiny wings from the lateral crown areas; a katoporodid− type histological structure distinguished by a network of branched wide dentine canals. Other scales with a notch on a smooth rhomboidal crown and postero−laterally down−stepped lateral rims have many characters in common with Loganellia grossi. Associated with the thelodonts are indeterminable acanthodian scales and a possible dentigerous jaw bone fragment. This finding also provides evidence of a hitherto unknown southward dispersal of Loganellia to the shelves of peri−Gondwana. Key words: Thelodonti, Phlebolepidiformes, Loganelliiformes, palaeobiogeography, Silurian, Niur Formation, Iran. Vachik Hairapetian [[email protected]], Department of Geology, Islamic Azad University, Khorasgan branch, PO Box 81595−158, Esfahan, Iran; Henning Blom [[email protected]], Subdepartment of Evolutionary Organismal Biology, Department of Physiol− ogy and Developmental Biology, Uppsala University, Norbyvägen 18A, SE−752 36 Uppsala, Sweden; C. -

Upcoming Projects Infrastructure Construction Division About Bane NOR Bane NOR Is a State-Owned Company Respon- Sible for the National Railway Infrastructure
1 Upcoming projects Infrastructure Construction Division About Bane NOR Bane NOR is a state-owned company respon- sible for the national railway infrastructure. Our mission is to ensure accessible railway infra- structure and efficient and user-friendly ser- vices, including the development of hubs and goods terminals. The company’s main responsible are: • Planning, development, administration, operation and maintenance of the national railway network • Traffic management • Administration and development of railway property Bane NOR has approximately 4,500 employees and the head office is based in Oslo, Norway. All plans and figures in this folder are preliminary and may be subject for change. 3 Never has more money been invested in Norwegian railway infrastructure. The InterCity rollout as described in this folder consists of several projects. These investments create great value for all travelers. In the coming years, departures will be more frequent, with reduced travel time within the InterCity operating area. We are living in an exciting and changing infrastructure environment, with a high activity level. Over the next three years Bane NOR plans to introduce contracts relating to a large number of mega projects to the market. Investment will continue until the InterCity rollout is completed as planned in 2034. Additionally, Bane NOR plans together with The Norwegian Public Roads Administration, to build a safer and faster rail and road system between Arna and Stanghelle on the Bergen Line (western part of Norway). We rely on close -

Fins, Limbs, and Tails: Outgrowths and Axial Patterning in Vertebrate Evolution Michael I
Review articles Fins, limbs, and tails: outgrowths and axial patterning in vertebrate evolution Michael I. Coates1* and Martin J. Cohn2 Summary Current phylogenies show that paired fins and limbs are unique to jawed verte- brates and their immediate ancestry. Such fins evolved first as a single pair extending from an anterior location, and later stabilized as two pairs at pectoral and pelvic levels. Fin number, identity, and position are therefore key issues in vertebrate developmental evolution. Localization of the AP levels at which develop- mental signals initiate outgrowth from the body wall may be determined by Hox gene expression patterns along the lateral plate mesoderm. This regionalization appears to be regulated independently of that in the paraxial mesoderm and axial skeleton. When combined with current hypotheses of Hox gene phylogenetic and functional diversity, these data suggest a new model of fin/limb developmental evolution. This coordinates body wall regions of outgrowth with primitive bound- aries established in the gut, as well as the fundamental nonequivalence of pectoral and pelvic structures. BioEssays 20:371–381, 1998. 1998 John Wiley & Sons, Inc. Introduction over and again to exemplify fundamental concepts in biological Vertebrate appendages include an amazing diversity of form, theory. The striking uniformity of teleost pectoral fin skeletons from the huge wing-like fins of manta rays or the stumpy limbs of illustrated Geoffroy Saint-Hilair’s discussion of ‘‘special analo- frogfishes, to ichthyosaur paddles, the extraordinary fingers of gies,’’1 while tetrapod limbs exemplified Owen’s2 related concept aye-ayes, and the fin-like wings of penguins. The functional of ‘‘homology’’; Darwin3 then employed precisely the same ex- diversity of these appendages is similarly vast and, in addition to ample as evidence of evolutionary descent from common ances- various modes of locomotion, fins and limbs are also used for try.