Micronesian Mangrove Forest Structure and Tree Responses to a Severe Typhoon
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A Summary of Palau's Typhoon History 1945-2013
A Summary of Palau’s Typhoon History 1945-2013 Coral Reef Research Foundation, Palau Dec, 2014 © Coral Reef Research Foundation 2014 Suggested citation: Coral Reef Research Foundation, 2014. A Summary of Palau’s Typhoon History. Technical Report, 17pp. www.coralreefpalau.org Additions and suggestions welcome. Please email: [email protected] 2 Summary: Since 1945 Palau has had 68 recorded typhoons, tropical storms or tropical depressions come within 200 nmi of its islands or reefs. At their nearest point to Palau, 20 of these were typhoon strength with winds ≥64kts, or an average of 1 typhoon every 3 years. November and December had the highest number of significant storms; July had none over 40 kts and August had no recorded storms. Data Compilation: Storms within 200 nmi (nautical miles) of Palau were identified from the Digital Typhoon, National Institute of Informatics, Japan web site (http://agora.ex.nii.ac.jp/digital- typhoon/reference/besttrack.html.en). The storm tracks and intensities were then obtained from the Joint Typhoon Warning Center (JTWC) (https://metoc.ndbc.noaa.gov/en/JTWC/). Three storm categories were used following the JTWC: Tropical Depression, winds ≤ 33 kts; Tropical Storm, winds 34-63 kts; Typhoon ≥64kts. All track data was from the JTWC archives. Tracks were plotted on Google Earth and the nearest distance to land or reef, and bearing from Palau, were measured; maximum sustained wind speed in knots (nautical miles/hr) at that point was recorded. Typhoon names were taken from the Digital Typhoon site, but typhoon numbers for the same typhoon were from the JTWC archives. -

Introduction to Emergency Management Second Edition
Introduction to Emergency Management Second Edition Introduction to Emergency Management Second Edition George D. Haddow Jane A. Bullock With Contributions by Damon P. Coppola Amsterdam • Boston • Heidelberg • London • New York • Oxford • Paris San Diego • San Francisco • Singapore • Sydney • Tokyo Elsevier Butterworth–Heinemann 30 Corporate Drive, Suite 400, Burlington, MA 01803, USA Linacre House, Jordan Hill, Oxford OX2 8DP, UK Copyright © 2006, Elsevier Inc. All rights reserved. No part of this publication may be reproduced, stored in a retrieval system, or transmitted in any form or by any means, electronic, mechanical, photocopying, recording, or otherwise, without the prior written permission of the publisher. Permissions may be sought directly from Elsevier’s Science & Technology Rights Department in Oxford, UK: phone: (+44) 1865 843830, fax: (+44) 1865 853333, e-mail: permissions@elsevier. co.uk. You may also complete your request on-line via the Elsevier homepage (http://elsevier.com), by selecting “Customer Support” and then “Obtaining Permissions.” Recognizing the importance of preserving what has been written, Elsevier prints its books on acid-free paper whenever possible. Library of Congress Cataloging-in-Publication Data Haddow, George D. Introduction to emergency management / George D. Haddow, Jane A. Bullock. — 2nd ed. p. cm. Includes bibliographical references and index. ISBN 0-7506-7961-1 (hardcover : alk. paper) 1. Emergency management. 2. Emergency management—United States. I. Bullock, Jane A. II. Title. HV551.2.H3 -

Ingredients for a Tropical Cyclone /Typhoon
Saffir-Simpson Tropical Cyclone Scale Examples of Satellite Imagery for Potential Damages Examples of Damage for Each (STCS) Each Category (structures, infrastructure, and vegetation) Category Tropical Storm Category A STCS is a scale that relates maximum tropical cyclone Damage only to the flimsiest lean-to type structure. TS CAT B TY CAT 1 Tropical cyclone is a general term that refers to Minor damage to banana and papaya trees. surface winds to potential damage and storm surge. A tropical depressions, tropical storms, and typhoons. similar scale has been used in the Atlantic for three decades. Guard and Lander (1999) adapted the Tropical Storm Category B TS CAT A TS CAT B Atlantic scale to tropical regions and added a tropical Major damage to huts made of thatch or loosely attached storm scale. STCS considers: tropical building corrugated sheet metal or plywood; sheet metal and materials/practices; tropical vegetation/agriculture; plywood may become airborne. Minor damage to weakening effects of termites, wood rot, salt spray; buildings of light materials. Moderate damage to banana and effects of the coral reefs on storm surge. and papaya trees. Damage to banana trees Weakened, unguyed Two tropical storm categories; five typhoon categories Typhoon Category 1 wooden power poles Corrugated metal and plywood stripped from poorly can break Note: Categories are based on 1-min. average maximum constructed or termite infested structures and may become sustained winds (MSW) and it’s corresponding 1-3 airborne. Few wooden, non-reinforced power poles tilted TY CAT 2 TY CAT 3 second peak gust. and some rotten power poles broken. -

The Causes of Mangrove Death on Yap, Palau, Pohnpei and Kosrae
Chapter II The Causes of Mangrove Death on Yap, Palau, Pohnpei, and Kosrae By: Phil G. Cannon, Margie Falanruw, Francis Ruegorong, Rich MacKenzie, Katie Friday, Amy L. Ross- Davis, Sara M. Ashiglar, Ned B. Klopfenstein; with Zhangfeng Liu, Mohammad Golabi, and Chancy Thomas Iyekar. Abstract: The area of a massive mangrove dieback in Yinuf Mn Island, Yap, was selected as the first location to study mangrove dieback problems. Seawater and soil samples were collected from plots where the mangrove trees were dead/dying and these samples were analyzed for eight different seawater and soil floor properties. Seawater and soil properties from dead/dying/missing mangrove samples were then compared with these same eight properties for seawater and soil samples collected in nearby plots where mangrove trees appeared healthy. For these eight properties, no apparent differences were found that might help explain why mangroves had died or stayed healthy. Consequently, other possible explanations for mangrove dieback were sought, based on current stand conditions and a consideration of extenuating circumstances in the recent history of this site. At this Yinuf Mn island site, a plausible explanation for the massive mangrove dieback was that Typhoon Sudal, which had struck 9 years earlier (in April, 2004) with both strong winds and a powerful storm surge, had pushed over about 100 acres (40.5 ha) of trees on the seaward edge of this mangrove stand. These trees may also have been heavily butt rotted or impacted by an oil spill, factors which could have rendered trees more prone to toppling during Typhoon Sudal. In a separate incident, Typhoon Bopha (December, 2012), caused minor but still measurable damage to a mature mangrove stand in Babeldaob, Palau. -

Hawaii and the US-Affiliated Pacific Islands. In
Hawaii and the U.S.-Affiliated Pacific Islands J.B. Friday, Kathleen Friday, and Craig Elevitch J.B. Friday is the extension forester, University of Hawaii at Manoa, College of Tropical Agriculture and Human Resources Cooperative Extension Service; Kathleen Friday is the Forest Legacy/Stewardship program manager—Hawai’i and Pacific Islands, U.S. Department of Agriculture, Forest Service, Pacific Southwest Region, State and Private Forestry; Craig Elevitch is the director of Agroforestry Net. Description of the Region systems. Continuous soil cover prevents erosion. Because the species diversity and structure of tree-based multistory gardens Hawaii and the U.S.-affiliated Pacific Islands (fig. A.1) include are similar to native forests, these agroforests protect water- a diversity of traditional and modern agroforestry systems that sheds and water quality, both in streams and near the shore. have developed across a broad range of environments, from The productivity over extremely long timeframes based only low coral atolls to high volcanic islands rising to 4,205 meters on local resources attests to their value as models for modern (m) (13,796 feet [ft]) in Hawaii. The peoples of Micronesia agroforestry systems that can be resilient to environmental and Polynesia settled their islands as many as 4,000 years ago stressors of the type that are projected to accompany climate (Athens and Ward 2004) and brought with them a basic suite change (table A.1). of agricultural plants. In the ensuing centuries, they developed highly sophisticated agroforestry systems tailored to meet food The most common traditional system is a tree-based multistory security needs within the local environments they inhabited. -

Federated States of Micronesiasidebar Federated States of Micronesia A
The State of Coral Reef Ecosystems of the Federated States of Micronesia Mike Hasurmai1, Eugene Joseph2, Steve Palik3, Kerat Rikim4 INTRODUCTION AND SETTING From east to west, the Freely Associated States include the Republic of the Marshall Islands (RMI), the Feder- Sidebar ated States of Micronesia (FSM), and the Republic of Palau. The FSM and Palau are known as the Caroline Islands, which span 2,500 km and are among the longest island chains in the world. All of these Micronesian islands were formerly a part of the Trust Territory of the Pacific Islands administered by the United States af- ter World War II (WWII). All three countries achieved independence within the past 25 years but retain close economic and strategic ties to the U.S. (Hezel, 1995). Although the process was initiated as early as 1979, the Compact of Free Association between the U.S. and the RMI and FSM did not go into effect until 1986; the Compact of Free Association between the U.S. and the Republic of Guam was effective in 1995. The FSM is comprised of four states, from east to west; Kosrae, Pohnpei, Chuuk, and Yap (Figure 14.1). Each island or group has its own language, customs, local government, and traditional system for managing marine resources. The FSM has both high islands and atolls, and islanders have a strong dependence on coral reefs and marine resources, both economically and culturally. The islands support three basic reef formations: fringing reefs, barrier reefs, and atolls, which correspond to the stage of reef development at each island. -

Biological Control of Chromolaena Odorata in the American Pacific Micronesian Islands
49 Biological control of Chromolaena odorata in the American Pacific Micronesian Islands R. Muniappanl, K. Englberger2 and G.V.P. Reddy3 1IPM CRSP, OIRED, Virginia Tech, 526 Prices Fork Road, Blacksburg, VA 24061, Email: [email protected] 2Coordinator, Plant Protection Micronesia, Secretariat of the Pacific Community, P.O. Box 2299, Palikir, Pohnpei 96941, Federated States of Micronesia 3Western Pacific Tropical Research Center, College of Natural and Applied Sciences, University of Guam, Mangilao, Guam 96923, USA Abstract. The program for biological control of Chromolaena odorata in the American Pacific Islands in Micronesia was supported by the TSTAR program of CSREES, USDA. The natural enemy, an arctiid caterpillar, Pareuchaetes pseudoinsulata has established in Guam, Rota, Saipan, and Tinian in the Mariana Islands, Chuuk, Kosrae and Pohnpei in the East Caroline Islands and Yap in the West Caroline Islands. The second natural enemy, the gall fly, Cecidochares connexa has established in all the Mariana and Caroline Islands. The eriophyid mite, Acalitus adoratus has fortuitously established in all the Mariana and Caroline Islands. Chromolaena has been suppressed and it is no longer a concern in the islands wherein P. pseudoinsulata and C. connexa have already been established. An eradication program is in progress in the Marshall Islands. Introduction The island groups Marianas, Carolines and Marshalls constitute the Micronesian Islands associated with the United States of America. They comprise of both coral atolls and volcanic islands. Subsistence agriculture is the most prevalent operation with a few commercial farming activities. The invasive weed, Chromolaena odorata (L.) King and Robinson (Asteraceae), first observed in the early 1960s in Guam has subsequently spread to the rest of the American Pacific Micronesian Islands. -

FEMA Disaster Cost-Shares: Evolution and Analysis
FEMA Disaster Cost-Shares: Evolution and Analysis Francis X. McCarthy Analyst in Emergency Management Policy March 9, 2010 Congressional Research Service 7-5700 www.crs.gov R41101 CRS Report for Congress Prepared for Members and Committees of Congress FEMA Disaster Cost-Shares: Evolution and Analysis Summary The Robert T. Stafford Disaster Relief and Emergency Assistance Act (The Stafford Act, P.L. 93- 288) contains discretion for the President to adjust cost-shares for the Public Assistance (PA) program, Sections 406 and 407 of the act, that provides assistance to states, local governments and non-profit organizations for debris removal and rebuilding of the public and non-profit infrastructure. The language of the Stafford Act defining cost-shares for the repair, restoration, and replacement of damaged facilities provides that the federal share “shall be not less than 75 percent.” These provisions have been in effect for over 20 years. While the authority to adjust the cost-share is long standing, the history of FEMA’s administrative adjustments and Congress’ legislative actions in this area, are of a more recent vintage. In all, there have been 222 cost-share adjustments of varying sizes and lengths of time. In 1998 FEMA promulgated, in regulation, a more consistent and open approach to cost-share adjustments. The overwhelming majority of these actions have been based on that regulatory authority and carried out by the executive branch through administrative actions. However, since 1997, and particularly in the wake of the difficult issues caused by the Gulf Coast storms of 2005, Congress has begun to exercise its authority to adjust cost-shares. -

Micronesia to the UN 820 2Nd Avenue, Suite 17A Telephone: (212) 697-8370 New York, N.Y
Permanent Mission of the Federated States of Micronesia to the UN 820 2nd Avenue, Suite 17A Telephone: (212) 697-8370 New York, N.Y. 10017 Facsimile: (212) 697-8295 e-mail: [email protected] http://www.fsmgov.org/ ______________________________________________________________________________________________________________________________________________________________________________________________________________________________________________ Federated States of Micronesia Views on the Possible Security Implications of Climate Change to be included in the report of the Secretary-General to the 64 th Session of the United Nations General Assembly Submitted pursuant to UN General Assembly Resolution A/RES/63/281 Introduction On June 3, 2009, the United Nations General Assembly unanimously adopted Resolution A/RES/63/281 inviting the relevant organs of the United Nations to intensify their efforts in considering and addressing climate change, especially its security implications. It represents the first time that the security implications of climate change have been brought to the attention of the international community. Under the terms of operative paragraph 2, the resolution “requests the Secretary-General to submit a comprehensive report to the General Assembly at its sixty-fourth session on the possible security implications of climate change, based on the views of the Member States and relevant regional and international organizations.” The Federated States of Micronesia (FSM) fully aligns itself with the submission by the Pacific Small Island Developing States made by Nauru and hereby provides additional views and input for the above-mentioned report. General description of the Federated States of Micronesia With its Environmental Vulnerability Index at 392, 1 the future security of the Federated States of Micronesia (FSM) is at high risk from the adverse impacts of climate change. -

Effects of Super Typhoons on Cyclonic Ocean Eddies in the Western North
PUBLICATIONS Journal of Geophysical Research: Oceans AUTHOR’S PREFACE Effects of super typhoons on cyclonic ocean eddies in the TO A SPECIAL western North Pacific: A satellite data-based evaluation COLLECTION 10.1002/2013JC009575 between 2000 and 2008 Liang Sun1,2, Ying-Xin Li1, Yuan-Jian Yang1,3, Qiaoyan Wu2, Xue-Tao Chen1, Qiu-Yang Li1, Yu-Bin Li4, Special Section: and Tao Xian1 Western Pacific Ocean Circulation and Climate 1Key Laboratory of the Atmospheric Composition and Optical Radiation, CAS, School of Earth and Space Sciences, University of Science and Technology of China, Hefei, People’s Republic of China, 2State Key Laboratory of Satellite Ocean Correspondence to: Environment Dynamics, Second Institute of Oceanography, State Oceanic Administration, Hangzhou, People’s Republic of L. Sun, 3 [email protected] China, Key Laboratory of Atmospheric Sciences and Satellite Remote Sensing of Anhui Province, Anhui Institute of Meteorological Sciences, Hefei, People’s Republic of China, 4School of Applied Meteorology, Nanjing University of Information Science and Technology, Nanjing, People’s Republic of China Citation: Sun, L., Y.-X. Li, Y.-J. Yang, Q. Wu, X.-T. Chen, Q.-Y. Li, Y.-B. Li, and T. Xian (2014), Effects of super typhoons on Abstract A composite time series of the merged satellite altimeters sea surface height anomaly (SSHA) cyclonic ocean eddies in the western data and satellite-observed sea surface temperature (SST) data were used to identify eddies in the Western North Pacific: A satellite data-based North Pacific Ocean (WNPO), where there were numbers of intense typhoons. This study systematically evaluation between 2000 and 2008, J. -
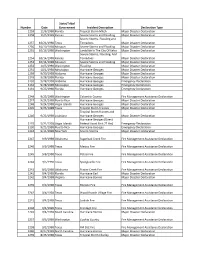
Identification of Disaster Code Declaration
State/Tribal Number Date Government Incident Description Declaration Type 1259 11/6/1998 Florida Tropical Storm Mitch Major Disaster Declaration 1258 11/5/1998 Kansas Severe Storms and Flooding Major Disaster Declaration Severe Storms, Flooding and 1257 10/21/1998 Texas Tornadoes Major Disaster Declaration 1256 10/19/1998 Missouri Severe Storms and Flooding Major Disaster Declaration 1255 10/16/1998 Washington Landslide In The City Of Kelso Major Disaster Declaration Severe Storms, Flooding, And 1254 10/14/1998 Kansas Tornadoes Major Disaster Declaration 1253 10/14/1998 Missouri Severe Storms and Flooding Major Disaster Declaration 1252 10/5/1998 Washington Flooding Major Disaster Declaration 1251 10/1/1998 Mississippi Hurricane Georges Major Disaster Declaration 1250 9/30/1998 Alabama Hurricane Georges Major Disaster Declaration 1249 9/28/1998 Florida Hurricane Georges Major Disaster Declaration 3133 9/28/1998 Alabama Hurricane Georges Emergency Declaration 3132 9/28/1998 Mississippi Hurricane Georges Emergency Declaration 3131 9/25/1998 Florida Hurricane Georges Emergency Declaration 2248 9/25/1998 Washington Columbia County Fire Management Assistance Declaration 1247 9/24/1998 Puerto Rico Hurricane Georges Major Disaster Declaration 1248 9/24/1998 Virgin Islands Hurricane Georges Major Disaster Declaration 1245 9/23/1998 Texas Tropical Storm Frances Major Disaster Declaration Tropical Storm Frances and 1246 9/23/1998 Louisiana Hurricane Georges Major Disaster Declaration Hurricane Georges (Direct 3129 9/21/1998 Virgin Islands Federal -

4.7. Predicting Tropical Cyclone Intensification
The Association of Tall Eyewall Convection with Tropical Cyclone Intensification A dissertation submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy at George Mason University By Owen A. Kelley Master of Science George Mason University, 1997 Bachelor of Arts St. John's College, 1993 Director: Michael Summers, Professor Department of Physics and Astronomy Spring Semester 2008 George Mason University Fairfax, VA Copyright 2008 Owen A. Kelley ii ACKNOWLEDGEMENTS Over the past four years, a number of people have discussed with me the ideas in this dissertation. They include Tom Bell, Natalie Blades, Craig Bohren, Scott Braun, Daniel Cecil, Jeff Halverson, Bart Kelley, Genevieve Demos Kelley, John Kwiatkowski, Chris Landsea, Chuntao Liu, David Marks, Daniel Melendez, Mike McCumber, Bob Meneghini, Bill Olson, Dave Silberstein, Joanne Simpson, Erich Stocker, John Stout, and Ed Zipser. The above mentioned people do not necessarily agree with the conclusions of this dissertation, nor are they responsible for any errors. Section 3.9 contains unpublished research from a collaboration with John Stout. The TRMM Science Data and Information System (TSDIS) provided computational facilities. The dissertation was strengthened by suggestions from my adviser Michael Summers and my dissertation committee, which includes Jim Beall, Jeff Halverson, and Menas Kafatos. Various organizations and individuals provided the data used in this dissertation. NASA/JAXA/NICT provided TRMM data. Many TRMM subsets were downloaded from the JAXA Tropical Cyclone Database. NCDC provided ground radar data. The National Hurricane Center (NHC) and the Joint Typhoon Warning Center (JTWC) provided world-wide best track wind intensity estimates. Scott Braun provided output from his simulation of Hurricane Bonnie (1998).