Acetylcholine Induces Yeast to Hyphal Form Transition in Candida Albicans
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Alzheimer's Disease Protective P522R Variant of PLCG2
bioRxiv preprint doi: https://doi.org/10.1101/2020.04.27.059600; this version posted April 28, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY 4.0 International license. The Alzheimer’s disease protective P522R variant of PLCG2, consistently enhances stimulus-dependent PLCγ2 activation, depleting substrate and altering cell function. Emily Maguire #1, Georgina E. Menzies#1, Thomas Phillips#1, Michael Sasner2, Harriet M. Williams2, Magdalena A. Czubala3, Neil Evans1, Emma L Cope4, Rebecca Sims5, Gareth R. Howell2, Emyr Lloyd-Evans4, Julie Williams†1,5, Nicholas D. Allen†4 and Philip R. Taylor†*1,3. 1 UK Dementia Research Institute at Cardiff, Hadyn Ellis Building, Maindy Road, Cardiff, CF24 4HQ, Wales, UK. 2 The Jackson Laboratory, Bar Harbor, Maine 04660, USA. 3 Systems Immunity University Research Institute, Tenovus Building, Heath Park, Cardiff CF 14 4XN, Wales, UK. 4 School of Biosciences, Cardiff University, Museum Avenue, Cardiff, CF10 3AX. 5 MRC Centre for Neuropsychiatric Genetics & Genomics, Hadyn Ellis Building, Maindy Road, Cardiff, CF24 4HQ, Wales, UK. #†These authors contributed equally *To whom correspondence should be addressed (lead contact): Prof Philip R. Taylor; Tel: +44(0)2920687328; Email: [email protected]. Abstract: Recent genome-wide association studies of Alzheimer’s disease (AD) have identified variants implicating immune pathways in disease development. A rare coding variant of PLCG2, which encodes PLCγ2, shows a significant protective effect for AD (rs72824905, P522R, P=5.38x10-10, Odds Ratio = 0.68). -

Supplemental Figure 1. Vimentin
Double mutant specific genes Transcript gene_assignment Gene Symbol RefSeq FDR Fold- FDR Fold- FDR Fold- ID (single vs. Change (double Change (double Change wt) (single vs. wt) (double vs. single) (double vs. wt) vs. wt) vs. single) 10485013 BC085239 // 1110051M20Rik // RIKEN cDNA 1110051M20 gene // 2 E1 // 228356 /// NM 1110051M20Ri BC085239 0.164013 -1.38517 0.0345128 -2.24228 0.154535 -1.61877 k 10358717 NM_197990 // 1700025G04Rik // RIKEN cDNA 1700025G04 gene // 1 G2 // 69399 /// BC 1700025G04Rik NM_197990 0.142593 -1.37878 0.0212926 -3.13385 0.093068 -2.27291 10358713 NM_197990 // 1700025G04Rik // RIKEN cDNA 1700025G04 gene // 1 G2 // 69399 1700025G04Rik NM_197990 0.0655213 -1.71563 0.0222468 -2.32498 0.166843 -1.35517 10481312 NM_027283 // 1700026L06Rik // RIKEN cDNA 1700026L06 gene // 2 A3 // 69987 /// EN 1700026L06Rik NM_027283 0.0503754 -1.46385 0.0140999 -2.19537 0.0825609 -1.49972 10351465 BC150846 // 1700084C01Rik // RIKEN cDNA 1700084C01 gene // 1 H3 // 78465 /// NM_ 1700084C01Rik BC150846 0.107391 -1.5916 0.0385418 -2.05801 0.295457 -1.29305 10569654 AK007416 // 1810010D01Rik // RIKEN cDNA 1810010D01 gene // 7 F5 // 381935 /// XR 1810010D01Rik AK007416 0.145576 1.69432 0.0476957 2.51662 0.288571 1.48533 10508883 NM_001083916 // 1810019J16Rik // RIKEN cDNA 1810019J16 gene // 4 D2.3 // 69073 / 1810019J16Rik NM_001083916 0.0533206 1.57139 0.0145433 2.56417 0.0836674 1.63179 10585282 ENSMUST00000050829 // 2010007H06Rik // RIKEN cDNA 2010007H06 gene // --- // 6984 2010007H06Rik ENSMUST00000050829 0.129914 -1.71998 0.0434862 -2.51672 -

Genazzani-2003-Calcineurin And
Edinburgh Research Explorer Calcineurin controls the expression of numerous genes in cerebellar granule cells Citation for published version: Kramer, D, Fresu, L, Ashby, DS, Freeman, TC & Genazzani, AA 2003, 'Calcineurin controls the expression of numerous genes in cerebellar granule cells', Molecular and Cellular Neuroscience, vol. 23, no. 2, pp. 325- 30. Link: Link to publication record in Edinburgh Research Explorer Document Version: Publisher's PDF, also known as Version of record Published In: Molecular and Cellular Neuroscience Publisher Rights Statement: © 2003 Elsevier Science (USA) General rights Copyright for the publications made accessible via the Edinburgh Research Explorer is retained by the author(s) and / or other copyright owners and it is a condition of accessing these publications that users recognise and abide by the legal requirements associated with these rights. Take down policy The University of Edinburgh has made every reasonable effort to ensure that Edinburgh Research Explorer content complies with UK legislation. If you believe that the public display of this file breaches copyright please contact [email protected] providing details, and we will remove access to the work immediately and investigate your claim. Download date: 28. Sep. 2021 Molecular and Cellular Neuroscience 23 (2003) 325–330 www.elsevier.com/locate/ymcne Calcineurin controls the expression of numerous genes in cerebellar granule cells Dana Kramer,a Luigia Fresu,b Dominique S. Ashby,a Tom C. Freeman,c and Armando A. Genazzania,* a Department of Pharmacology, Tennis Court Road, Cambridge, CB2 1PD, UK b Department of Pharmacology, Gynaecology and Obstetrics, University of Sassari, Italy c Microarray Group, MRC HGMP-RC, Wellcome Trust Genome Campus, Hinxton, CB10 1SB, UK Received 31 October 2002; revised 7 January 2003; accepted 20 January 2003 Abstract The Ca2ϩ/calmodulin-dependent phosphatase calcineurin plays a crucial role in gene expression in different cell types such as T-lymphocytes, cardiac myocytes, and smooth muscle cells. -

Viewed Under 23 (B) Or 203 (C) fi M M Male Cko Mice, and Largely Unaffected Magni Cation; Scale Bars, 500 M (B) and 50 M (C)
BRIEF COMMUNICATION www.jasn.org Renal Fanconi Syndrome and Hypophosphatemic Rickets in the Absence of Xenotropic and Polytropic Retroviral Receptor in the Nephron Camille Ansermet,* Matthias B. Moor,* Gabriel Centeno,* Muriel Auberson,* † † ‡ Dorothy Zhang Hu, Roland Baron, Svetlana Nikolaeva,* Barbara Haenzi,* | Natalya Katanaeva,* Ivan Gautschi,* Vladimir Katanaev,*§ Samuel Rotman, Robert Koesters,¶ †† Laurent Schild,* Sylvain Pradervand,** Olivier Bonny,* and Dmitri Firsov* BRIEF COMMUNICATION *Department of Pharmacology and Toxicology and **Genomic Technologies Facility, University of Lausanne, Lausanne, Switzerland; †Department of Oral Medicine, Infection, and Immunity, Harvard School of Dental Medicine, Boston, Massachusetts; ‡Institute of Evolutionary Physiology and Biochemistry, St. Petersburg, Russia; §School of Biomedicine, Far Eastern Federal University, Vladivostok, Russia; |Services of Pathology and ††Nephrology, Department of Medicine, University Hospital of Lausanne, Lausanne, Switzerland; and ¶Université Pierre et Marie Curie, Paris, France ABSTRACT Tight control of extracellular and intracellular inorganic phosphate (Pi) levels is crit- leaves.4 Most recently, Legati et al. have ical to most biochemical and physiologic processes. Urinary Pi is freely filtered at the shown an association between genetic kidney glomerulus and is reabsorbed in the renal tubule by the action of the apical polymorphisms in Xpr1 and primary fa- sodium-dependent phosphate transporters, NaPi-IIa/NaPi-IIc/Pit2. However, the milial brain calcification disorder.5 How- molecular identity of the protein(s) participating in the basolateral Pi efflux remains ever, the role of XPR1 in the maintenance unknown. Evidence has suggested that xenotropic and polytropic retroviral recep- of Pi homeostasis remains unknown. Here, tor 1 (XPR1) might be involved in this process. Here, we show that conditional in- we addressed this issue in mice deficient for activation of Xpr1 in the renal tubule in mice resulted in impaired renal Pi Xpr1 in the nephron. -

Silencing of Phosphoinositide-Specific
ANTICANCER RESEARCH 34: 4069-4076 (2014) Silencing of Phosphoinositide-specific Phospholipase C ε Remodulates the Expression of the Phosphoinositide Signal Transduction Pathway in Human Osteosarcoma Cell Lines VINCENZA RITA LO VASCO1, MARTINA LEOPIZZI2, DANIELA STOPPOLONI3 and CARLO DELLA ROCCA2 Departments of 1Sense Organs , 2Medicine and Surgery Sciences and Biotechnologies and 3Biochemistry Sciences “A. Rossi Fanelli”, Sapienza University, Rome, Italy Abstract. Background: Ezrin, a member of the signal transduction pathway (5). The reduction of PIP2 ezrin–radixin–moesin family, is involved in the metastatic induces ezrin dissociation from the plasma membrane (6). spread of osteosarcoma. Ezrin binds phosphatydil inositol-4,5- The levels of PIP2 are regulated by the PI-specific bisphosphate (PIP2), a crucial molecule of the phospholipase C (PI-PLC) family (7), constituting thirteen phosphoinositide signal transduction pathway. PIP2 levels are enzymes divided into six sub-families on the basis of amino regulated by phosphoinositide-specific phospholipase C (PI- acid sequence, domain structure, mechanism of recruitment PLC) enzymes. PI-PLCε isoform, a well-characterized direct and tissue distribution (7-15). PI-PLCε, a direct effector of effector of rat sarcoma (RAS), is at a unique convergence RAS (14-15), might be the point of convergence for the point for the broad range of signaling pathways that promote broad range of signalling pathways that promote the RAS GTPase-mediated signalling. Materials and Methods. By RASGTPase-mediated signalling (16). using molecular biology methods and microscopic analyses, In previous studies, we suggested a relationship between we analyzed the expression of ezrin and PLC genes after PI-PLC expression and ezrin (17-18). -

Genetic Studies of Bipolar Disorder in Patients Selectedby Their Treatment
medigraphic Artemisaen línea Salud Mental 2008;31:431-440 Genetic studies of bipolar disorder Genetic studies of bipolar disorder in patients selected by their treatment response* Abigail Ortiz-Domínguez,1 Martin Alda1 Conferencia magistral SUMMARY rapid cycling and non-episodic course in the lamotrigine group) and co-morbidity, with the lamotrigine-responder group showing a higher Bipolar disorder (BD) is a major mood disorder with several genes frequency of panic attacks and substance abuse. of moderate or small effect contributing to the genetic susceptibility. In conclusion, pharmacogenetic studies may provide important It is also likely heterogeneous, which stimulated efforts to refine its clues to the nature of bipolar disorder and the response to long term clinical phenotype, studies investigating the link between BD treatment. susceptibility and response to a specific mood stabilizer appear to be one of the promising directions. Key words: Lithium response, pharmacogenetic, probands. In particular, excellent response to lithium prophylaxis has been described as a clinical marker of a more homogeneous subgroup of BD, characterized by an episodic course, low rates of co-morbid RESUMEN conditions, absence of rapid cycling, and a strong genetic loading. These results also suggest that lithium response clusters in families El trastorno bipolar (TB) es un trastorno afectivo con varios genes, de (independent of the increased familial loading for affective disorders), efecto leve o moderado, que contribuyen a su susceptibilidad. Es likely on a genetic basis. asimismo un trastorno heterogéneo, lo que ha estimulado diversas For almost 40 years, clinical studies have pointed to differences iniciativas para refinar el fenotipo de los pacientes con este trastor- between lithium responders (LR) and non-responders (LNR). -

Plasma Lipidome Is Dysregulated in Alzheimer's Disease and Is
Liu et al. Translational Psychiatry (2021) 11:344 https://doi.org/10.1038/s41398-021-01362-2 Translational Psychiatry ARTICLE Open Access Plasma lipidome is dysregulated in Alzheimer’s disease and is associated with disease risk genes Yue Liu1,2, Anbupalam Thalamuthu1, Karen A. Mather1,3, John Crawford1, Marina Ulanova1, Matthew Wai Kin Wong1, Russell Pickford4, Perminder S. Sachdev 1,5 and Nady Braidy 1,6 Abstract Lipidomics research could provide insights of pathobiological mechanisms in Alzheimer’s disease. This study explores a battery of plasma lipids that can differentiate Alzheimer’s disease (AD) patients from healthy controls and determines whether lipid profiles correlate with genetic risk for AD. AD plasma samples were collected from the Sydney Memory and Ageing Study (MAS) Sydney, Australia (aged range 75–97 years; 51.2% male). Untargeted lipidomics analysis was performed by liquid chromatography coupled–mass spectrometry (LC–MS/MS). We found that several lipid species from nine lipid classes, particularly sphingomyelins (SMs), cholesterol esters (ChEs), phosphatidylcholines (PCs), phosphatidylethanolamines (PIs), phosphatidylinositols (PIs), and triglycerides (TGs) are dysregulated in AD patients and may help discriminate them from healthy controls. However, when the lipid species were grouped together into lipid subgroups, only the DG group was significantly higher in AD. ChEs, SMs, and TGs resulted in good classification accuracy using the Glmnet algorithm (elastic net penalization for the generalized linear model [glm]) with more than 80% AUC. In general, group lipids and the lipid subclasses LPC and PE had less classification accuracy compared to the other subclasses. We also found significant increases in SMs, PIs, and the LPE/PE ratio in human U251 astroglioma cell lines exposed to pathophysiological concentrations of oligomeric Aβ42. -
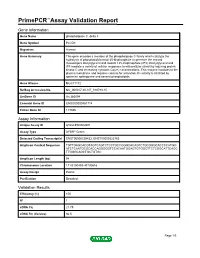
Primepcr™Assay Validation Report
PrimePCR™Assay Validation Report Gene Information Gene Name phospholipase C, delta 3 Gene Symbol PLCD3 Organism Human Gene Summary This gene encodes a member of the phospholipase C family which catalyze the hydrolysis of phosphatidylinositol 45-bisphosphate to generate the second messengers diacylglycerol and inositol 145-trisphosphate (IP3). Diacylglycerol and IP3 mediate a variety of cellular responses to extracellular stimuli by inducing protein kinase C and increasing cytosolic Ca(2+) concentrations. This enzyme localizes to the plasma membrane and requires calcium for activation. Its activity is inhibited by spermine sphingosine and several phospholipids. Gene Aliases MGC71172 RefSeq Accession No. NC_000017.10, NT_010783.15 UniGene ID Hs.380094 Ensembl Gene ID ENSG00000161714 Entrez Gene ID 113026 Assay Information Unique Assay ID qHsaCED0002601 Assay Type SYBR® Green Detected Coding Transcript(s) ENST00000539433, ENST00000322765 Amplicon Context Sequence TGTTGAGCACGTAGTCAGTCTCCTGCCGGGCACAGTCTGCGGGCACCCCATGG ATCTCAATGCGCACCAGGGGGTCCACAATGGAGTGTGGCTTCTCGGCATTCAGC TTGGGCAGCTGCTGTGC Amplicon Length (bp) 94 Chromosome Location 17:43190493-43190616 Assay Design Exonic Purification Desalted Validation Results Efficiency (%) 100 R2 1 cDNA Cq 21.75 cDNA Tm (Celsius) 86.5 Page 1/5 PrimePCR™Assay Validation Report gDNA Cq 23.62 Specificity (%) 100 Information to assist with data interpretation is provided at the end of this report. Page 2/5 PrimePCR™Assay Validation Report PLCD3, Human Amplification Plot Amplification of cDNA generated from -

Genetic Defect in Phospholipase Cd1 Protects Mice from Obesity By
ORIGINAL ARTICLE Genetic Defect in Phospholipase Cd1 Protects Mice From Obesity by Regulating Thermogenesis and Adipogenesis Masayuki Hirata,1 Mutsumi Suzuki,1 Rika Ishii,1 Reiko Satow,1 Takafumi Uchida,1 Tomoya Kitazumi,2 Tsutomu Sasaki,2 Tadahiro Kitamura,2 Hideki Yamaguchi,3,4 Yoshikazu Nakamura,1 and Kiyoko Fukami1 OBJECTIVE—Regulation of obesity development is an impor- size of adipocytes is a hallmark of obesity. The former tant issue to prevent metabolic syndromes. Gene-disrupted mice seems to be caused by proliferation and differentiation of of phospholipase Cd1(PLCd1), a key enzyme of phosphoinositide preadipocytes. On the other hand, the diet-induced in- turnover, seemed to show leanness. Here we examined whether crease in cell size is characterized by adipocyte hypertro- and how PLCd1 is involved in obesity. phy, which may be primarily caused by excessive lipid RESEARCH DESIGN AND METHODS—Weight gain, insulin overload and a decrease in metabolic rate. sensitivity, and metabolic rate in PLCd12/2 mice were compared Brown adipose tissue (BAT) is implicated in thermo- with PLCd1+/2 littermate mice on a high-fat diet. Thermogenic genesis and metabolic enhancement (5). Recent reports in- and adipogenetic potentials of PLCd12/2 immortalized brown dicated that BAT and skeletal muscle originate from adipocytes and adipogenesis of PLCd1-knockdown (KD) 3T3L1 a common precursor cell (6–9). Like skeletal muscle, BAT 2 2 cells, or PLCd1 / white adipose tissue (WAT) stromal-vascular plays a role in thermogenesis by promoting the expression of fraction (SVF) cells, were also investigated. a thermogenic gene, uncoupling protein 1 (UCP1). Upregu- RESULTS—PLCd12/2 mice showed marked decreases in weight lation of UCP1 by genetic manipulations or pharmacological gain and mass of epididymal WAT and preserved insulin sensitivity agents has been shown to reduce obesity and improve in- compared with PLCd1+/2 mice on a high-fat diet. -

Supplementary Table S4. FGA Co-Expressed Gene List in LUAD
Supplementary Table S4. FGA co-expressed gene list in LUAD tumors Symbol R Locus Description FGG 0.919 4q28 fibrinogen gamma chain FGL1 0.635 8p22 fibrinogen-like 1 SLC7A2 0.536 8p22 solute carrier family 7 (cationic amino acid transporter, y+ system), member 2 DUSP4 0.521 8p12-p11 dual specificity phosphatase 4 HAL 0.51 12q22-q24.1histidine ammonia-lyase PDE4D 0.499 5q12 phosphodiesterase 4D, cAMP-specific FURIN 0.497 15q26.1 furin (paired basic amino acid cleaving enzyme) CPS1 0.49 2q35 carbamoyl-phosphate synthase 1, mitochondrial TESC 0.478 12q24.22 tescalcin INHA 0.465 2q35 inhibin, alpha S100P 0.461 4p16 S100 calcium binding protein P VPS37A 0.447 8p22 vacuolar protein sorting 37 homolog A (S. cerevisiae) SLC16A14 0.447 2q36.3 solute carrier family 16, member 14 PPARGC1A 0.443 4p15.1 peroxisome proliferator-activated receptor gamma, coactivator 1 alpha SIK1 0.435 21q22.3 salt-inducible kinase 1 IRS2 0.434 13q34 insulin receptor substrate 2 RND1 0.433 12q12 Rho family GTPase 1 HGD 0.433 3q13.33 homogentisate 1,2-dioxygenase PTP4A1 0.432 6q12 protein tyrosine phosphatase type IVA, member 1 C8orf4 0.428 8p11.2 chromosome 8 open reading frame 4 DDC 0.427 7p12.2 dopa decarboxylase (aromatic L-amino acid decarboxylase) TACC2 0.427 10q26 transforming, acidic coiled-coil containing protein 2 MUC13 0.422 3q21.2 mucin 13, cell surface associated C5 0.412 9q33-q34 complement component 5 NR4A2 0.412 2q22-q23 nuclear receptor subfamily 4, group A, member 2 EYS 0.411 6q12 eyes shut homolog (Drosophila) GPX2 0.406 14q24.1 glutathione peroxidase -

Genetic Variants in PLCB4/PLCB1 As Susceptibility Loci for Coronary Artery
www.nature.com/scientificreports OPEN Genetic variants in PLCB4/PLCB1 as susceptibility loci for coronary artery aneurysm formation in Received: 27 March 2015 Accepted: 04 September 2015 Kawasaki disease in Han Chinese Published: 05 October 2015 in Taiwan Ying-Ju Lin1,2, Jeng-Sheng Chang3,4, Xiang Liu5, Hsinyi Tsang5, Wen-Kuei Chien6, Jin-Hua Chen6,7, Hsin-Yang Hsieh3,8, Kai-Chung Hsueh9, Yi-Tzone Shiao10, Ju-Pi Li2,11, Cheng-Wen Lin12, Chih-Ho Lai13, Jer-Yuarn Wu2,14, Chien-Hsiun Chen2,14, Jaung-Geng Lin2, Ting-Hsu Lin1, Chiu-Chu Liao1, Shao-Mei Huang1, Yu-Ching Lan15, Tsung-Jung Ho2, Wen-Miin Liang16, Yi-Chun Yeh16, Jung-Chun Lin17 & Fuu-Jen Tsai1,2,18 Kawasaki disease (KD) is an acute, inflammatory, and self-limited vasculitis affecting infants and young children. Coronary artery aneurysm (CAA) formation is the major complication of KD and the leading cause of acquired cardiovascular disease among children. To identify susceptible loci that might predispose patients with KD to CAA formation, a genome-wide association screen was performed in a Taiwanese KD cohort. Patients with both KD and CAA had longer fever duration and delayed intravenous immunoglobulin treatment time. After adjusting for these factors, 100 susceptibility loci were identified. Four genes were identified from a single cluster of 35 using the Ingenuity Pathway Analysis (IPA) Knowledge Base. Silencing KCNQ5, PLCB1, PLCB4, and PLCL1 inhibited the effect of lipopolysaccharide-induced endothelial cell inflammation with varying degrees of proinflammatory cytokine expression. PLCB1 showed the most significant inhibition. Endothelial cell inflammation was also inhibited by using a phospholipase C (PLC) inhibitor. -

(12) Patent Application Publication (10) Pub. No.: US 2003/0082511 A1 Brown Et Al
US 20030082511A1 (19) United States (12) Patent Application Publication (10) Pub. No.: US 2003/0082511 A1 Brown et al. (43) Pub. Date: May 1, 2003 (54) IDENTIFICATION OF MODULATORY Publication Classification MOLECULES USING INDUCIBLE PROMOTERS (51) Int. Cl." ............................... C12O 1/00; C12O 1/68 (52) U.S. Cl. ..................................................... 435/4; 435/6 (76) Inventors: Steven J. Brown, San Diego, CA (US); Damien J. Dunnington, San Diego, CA (US); Imran Clark, San Diego, CA (57) ABSTRACT (US) Correspondence Address: Methods for identifying an ion channel modulator, a target David B. Waller & Associates membrane receptor modulator molecule, and other modula 5677 Oberlin Drive tory molecules are disclosed, as well as cells and vectors for Suit 214 use in those methods. A polynucleotide encoding target is San Diego, CA 92121 (US) provided in a cell under control of an inducible promoter, and candidate modulatory molecules are contacted with the (21) Appl. No.: 09/965,201 cell after induction of the promoter to ascertain whether a change in a measurable physiological parameter occurs as a (22) Filed: Sep. 25, 2001 result of the candidate modulatory molecule. Patent Application Publication May 1, 2003 Sheet 1 of 8 US 2003/0082511 A1 KCNC1 cDNA F.G. 1 Patent Application Publication May 1, 2003 Sheet 2 of 8 US 2003/0082511 A1 49 - -9 G C EH H EH N t R M h so as se W M M MP N FIG.2 Patent Application Publication May 1, 2003 Sheet 3 of 8 US 2003/0082511 A1 FG. 3 Patent Application Publication May 1, 2003 Sheet 4 of 8 US 2003/0082511 A1 KCNC1 ITREXCHO KC 150 mM KC 2000000 so 100 mM induced Uninduced Steady state O 100 200 300 400 500 600 700 Time (seconds) FIG.