Annual Report 2009
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Taxa Named in Honor of Ihsan A. Al-Shehbaz
TAXA NAMED IN HONOR OF IHSAN A. AL-SHEHBAZ 1. Tribe Shehbazieae D. A. German, Turczaninowia 17(4): 22. 2014. 2. Shehbazia D. A. German, Turczaninowia 17(4): 20. 2014. 3. Shehbazia tibetica (Maxim.) D. A. German, Turczaninowia 17(4): 20. 2014. 4. Astragalus shehbazii Zarre & Podlech, Feddes Repert. 116: 70. 2005. 5. Bornmuellerantha alshehbaziana Dönmez & Mutlu, Novon 20: 265. 2010. 6. Centaurea shahbazii Ranjbar & Negaresh, Edinb. J. Bot. 71: 1. 2014. 7. Draba alshehbazii Klimeš & D. A. German, Bot. J. Linn. Soc. 158: 750. 2008. 8. Ferula shehbaziana S. A. Ahmad, Harvard Pap. Bot. 18: 99. 2013. 9. Matthiola shehbazii Ranjbar & Karami, Nordic J. Bot. doi: 10.1111/j.1756-1051.2013.00326.x, 10. Plocama alshehbazii F. O. Khass., D. Khamr., U. Khuzh. & Achilova, Stapfia 101: 25. 2014. 11. Alshehbazia Salariato & Zuloaga, Kew Bulletin …….. 2015 12. Alshehbzia hauthalii (Gilg & Muschl.) Salariato & Zuloaga 13. Ihsanalshehbazia Tahir Ali & Thines, Taxon 65: 93. 2016. 14. Ihsanalshehbazia granatensis (Boiss. & Reuter) Tahir Ali & Thines, Taxon 65. 93. 2016. 15. Aubrieta alshehbazii Dönmez, Uǧurlu & M.A.Koch, Phytotaxa 299. 104. 2017. 16. Silene shehbazii S.A.Ahmad, Novon 25: 131. 2017. PUBLICATIONS OF IHSAN A. AL-SHEHBAZ 1973 1. Al-Shehbaz, I. A. 1973. The biosystematics of the genus Thelypodium (Cruciferae). Contrib. Gray Herb. 204: 3-148. 1977 2. Al-Shehbaz, I. A. 1977. Protogyny, Cruciferae. Syst. Bot. 2: 327-333. 3. A. R. Al-Mayah & I. A. Al-Shehbaz. 1977. Chromosome numbers for some Leguminosae from Iraq. Bot. Notiser 130: 437-440. 1978 4. Al-Shehbaz, I. A. 1978. Chromosome number reports, certain Cruciferae from Iraq. -

A Study of the Floral Biology of Viciaria Amazonica (Poepp.) Sowerby (Nymphaeaceae)
A study of the Floral Biology of Viciaria amazonica (Poepp.) Sowerby (Nymphaeaceae) Ghillean T. Prance (1) Jorge R. Arias (2) Abstract Victoria and the beetles which visit the flowers in large numbers, and to collect data A field study of the floral biology of Victoria on V. amazonica to compare with the data of amazonica (Poepp.) Sowerby (Nymphaeaceae) was Valia & Girino (1972) on V. cruziana. made for comparison with the many studies made in cultivated plants, of Victoria in the past. In thE: study areas in the vicinity of Manaus, four species HISTORY OF WORK ON THE FLORAL of Dynastid beetles were found in flowers of V. BIOLOGY OF VICTORIA. amazonica, three of the genus Cyclocephala and one o! Ligyrus . The commonest species of beetle The nomenclatura( and taxonomic history proved to be a new species of Cyclocephala and was found in over 90 percent of the flowers studied. of the genus has already been summarized in The flowers of V. amazonica attract beetles by Prance (1974). where it has been shown that their odour and their white colour on the first the correct name for the Amazonian species day that they open. The beetles are trapped in the of Victoria is V. amazonica, and not the more flower for twenty-four hours and feed on the starchy carpellary appendages. Observations were frequently used name, V. regia. The taxonomic made of flower temperature, which is elevated up history is not treated further here. to 11 aC above ambient temperature, when the flower Victoria amazonica has been a subject of emits the odour to attract the beetles. -

Phylogenetic Position and Generic Limits of Arabidopsis (Brassicaceae)
PHYLOGENETIC POSITION Steve L. O'Kane, Jr.2 and Ihsan A. 3 AND GENERIC LIMITS OF Al-Shehbaz ARABIDOPSIS (BRASSICACEAE) BASED ON SEQUENCES OF NUCLEAR RIBOSOMAL DNA1 ABSTRACT The primary goals of this study were to assess the generic limits and monophyly of Arabidopsis and to investigate its relationships to related taxa in the family Brassicaceae. Sequences of the internal transcribed spacer region (ITS-1 and ITS-2) of nuclear ribosomal DNA, including 5.8S rDNA, were used in maximum parsimony analyses to construct phylogenetic trees. An attempt was made to include all species currently or recently included in Arabidopsis, as well as species suggested to be close relatives. Our ®ndings show that Arabidopsis, as traditionally recognized, is polyphyletic. The genus, as recircumscribed based on our results, (1) now includes species previously placed in Cardaminopsis and Hylandra as well as three species of Arabis and (2) excludes species now placed in Crucihimalaya, Beringia, Olimar- abidopsis, Pseudoarabidopsis, and Ianhedgea. Key words: Arabidopsis, Arabis, Beringia, Brassicaceae, Crucihimalaya, ITS phylogeny, Olimarabidopsis, Pseudoar- abidopsis. Arabidopsis thaliana (L.) Heynh. was ®rst rec- netic studies and has played a major role in un- ommended as a model plant for experimental ge- derstanding the various biological processes in netics over a half century ago (Laibach, 1943). In higher plants (see references in Somerville & Mey- recent years, many biologists worldwide have fo- erowitz, 2002). The intraspeci®c phylogeny of A. cused their research on this plant. As indicated by thaliana has been examined by Vander Zwan et al. Patrusky (1991), the widespread acceptance of A. (2000). Despite the acceptance of A. -

P020110307527551165137.Pdf
CONTENT 1.MESSAGE FROM DIRECTOR …………………………………………………………………………………………………………………………………………………… 03 2.ORGANIZATION STRUCTURE …………………………………………………………………………………………………………………………………………………… 05 3.HIGHLIGHTS OF ACHIEVEMENTS …………………………………………………………………………………………………………………………………………… 06 Coexistence of Conserve and Research----“The Germplasm Bank of Wild Species ” services biodiversity protection and socio-economic development ………………………………………………………………………………………………………………………………………………… 06 The Structure, Activity and New Drug Pre-Clinical Research of Monoterpene Indole Alkaloids ………………………………………… 09 Anti-Cancer Constituents in the Herb Medicine-Shengma (Cimicifuga L) ……………………………………………………………………………… 10 Floristic Study on the Seed Plants of Yaoshan Mountain in Northeast Yunnan …………………………………………………………………… 11 Higher Fungi Resources and Chemical Composition in Alpine and Sub-alpine Regions in Southwest China ……………………… 12 Research Progress on Natural Tobacco Mosaic Virus (TMV) Inhibitors…………………………………………………………………………………… 13 Predicting Global Change through Reconstruction Research of Paleoclimate………………………………………………………………………… 14 Chemical Composition of a traditional Chinese medicine-Swertia mileensis……………………………………………………………………………… 15 Mountain Ecosystem Research has Made New Progress ………………………………………………………………………………………………………… 16 Plant Cyclic Peptide has Made Important Progress ………………………………………………………………………………………………………………… 17 Progresses in Computational Chemistry Research ………………………………………………………………………………………………………………… 18 New Progress in the Total Synthesis of Natural Products ……………………………………………………………………………………………………… -
2279 Knapp-Checklisttag.Indd
A peer-reviewed open-access journal PhytoKeys 9: 15–179Checklist (2012) of vascular plants of the Department of Ñeembucú, Paraguay 15 doi: 10.3897/phytokeys.9.2279 CHECKLIST www.phytokeys.com Launched to accelerate biodiversity research Checklist of vascular plants of the Department of Ñeembucú, Paraguay Juana De Egea1,2, Maria Peña-Chocarro1, Cristina Espada1, Sandra Knapp1 1 Department of Botany, Th e Natural History Museum, Cromwell Road, London SW7 5BD, United Kingdom 2 Wildlife Conservation Society Paraguay, Capitán Benitez Vera 610, Asunción, Paraguay Corresponding author: S. Knapp ([email protected]) Academic editor: Susanne Renner | Received 25 October 2011 | Accepted 6 January 2012 | Published 30 January 2012 Citation: De Egea J, Peña-Chocarro M, Espada C, Knapp S (2012) Checklist of vascular plants of the Department of Ñeembucú, Paraguay. PhytoKeys 9: 15–179. doi: 10.3897/phytokeys.9.2279 Abstract Th e Department of Ñeembucú is one of the least well-documented areas of eastern Paraguay, and the fl ora is composed of a mixture of forest and Chaco elements. Regions like Ñeembucú are often considered of lower diversity and interest that more forested regions; this results from both actual species richness fi gures and from under-collecting due to perception as uninteresting. We present here a checklist of the vascular plants of Ñeembucú, which includes 676 taxa (including infraspecifi c taxa and collections identifi ed only to genus) in 100 families and 374 genera. Four hundred and thirty nine (439) of these are new records for Ñeembucú and of these, 4 are new published records for Paraguay. -

Pollen Ontogeny in Victoria (Nymphaeales)
Int. J. Plant Sci. 174(9):1259–1276. 2013. ᭧ 2013 by The University of Chicago. All rights reserved. 1058-5893/2013/17409-0006$15.00 DOI: 10.1086/673246 POLLEN ONTOGENY IN VICTORIA (NYMPHAEALES) Mackenzie L. Taylor,1,* Patrick J. Hudson,2,† Jolene M. Rigg,† Julie N. Strandquist,† Julie Schwartz Green,3,† Tara C. Thiemann,4,† and Jeffrey M. Osborn‡ *Department of Biology, Creighton University, Omaha, Nebraska 68178, USA; †Department of Biology, Truman State University, Kirksville, Missouri 63501, USA; and ‡School of Science, College of New Jersey, Ewing, New Jersey 08628, USA Editor: William E. Friedman Premise of research. Water lilies (Nymphaeales) make up one of the oldest independent lineages of an- giosperms. The giant water lily, Victoria, exhibits pollination and floral traits that are derived within Nym- phaeales. Specialization in pollination and floral biology is often reflected in pollen traits, and in Victoria,this is evidenced by the production of permanent tetrads. Compound pollen has evolved many times across the angiosperm phylogeny, but compound pollen development has been investigated in only a few taxa, and the degree of developmental variation in microspore cohesion is unknown. This article comprehensively charac- terizes the pollen ontogenetic sequence in Victoria for the first time. Methodology. Floral buds of Victoria amazonica, Victoria cruziana, and Longwood hybrid were field collected. Anthers at the sporogenous, microspore mother cell, tetrad, “free” microspore, and mature pollen grain stages were studied using combined LM/SEM/TEM. Pivotal results. Microspore cohesion in Victoria differs from that exhibited by the few compound pollen- producing taxa that have been studied. In Victoria, the calymmate tetrads fuse via crosswall cohesion, but cytoplasmic connections are transient and do not serve as a template for wall bridge formation. -

PART 2 AUTHOR INDEX of ARTICLES in the I.W.G.S. LIBRARY C = Copy Sheets
5 PART 2 AUTHOR INDEX OF ARTICLES IN THE I.W.G.S. LIBRARY c = copy sheets Abbott, Charles C. 1888. NYMPHAEA TUBEROSA IN EASTERN WATERS. Garden & Forest (issue unknown) c1 Ames, Oakes. 1900. AN INTERESTING GROUP OF NEW HYBRID BLOOMING NYMPHAEAS. American Gardening 21:644 c1 Anderson, Edgar. 1965. VICTORIA WATER LILIES. Mo. Bot. Gard. Bull. 53(5): 1-18 c11 Anderson, Fred. WATER LILIES FOR COOL SUMMER BEAUTY. Horticulture. August 1960: c2 Anderson, Michael G. & S.W. Idso. 1987. SURFACE GEOMETRY AND STOMATAL CONDUCTANCE EFFECTS ON EVAPORATION FROM AQUATIC MACROPHYTES. Water Resources Research. 23(6):1037-1042 c6 Andre {Editor of Revue Horticole}.1896. NEW HARDY WATER LILIES .The Garden 50:325 c1 Anthony, John. AN ILLUSTRATED LIFE OF SIR JOSEPH PAXTON. Shire Lifelines Book No. 21 c25 Armstrong, Wayne P. 1983a. A MARRIAGE BETWEEN A FERN AND AN ALGA. Environmental Southwest. Winter 1983: 20-24 c5 -----1983b THE WORLD'S SMALLEST WILDFLOWER. Environment Southwest, Summer 1983: 17-21 c5 Aston, Helen I. 1973. NYMPHOIDES OF AUSTRALIA. c13 -----1982 . NEW AUSTRALIAN SPECIES OF NYMPHOIDES. Muelleria 5:35-51 c17 -----1984. NYMPHOIDES TRIANGULARIS AND N. ELLIPTICA; TWO NEW AUSTRALIAN SPECIES. Muelleria 5265-270 c6 -----1985a. MONOCHORIA CYANEA AND M. AUSTRALASICA IN AUSTRALIA. Muelleria 651-57 c7. -----1985b. NYMPHOIDES OF PAPUA NEW GUINEA. Freshwater plants of PNG: 180-185 c6 -----1986. NYMPHOIDES DISPERMA. Muelleria 6(3):197-200 c4 -----1987a. LYMNOPHYTON AUSTRALIENSE A NEW GENERIC RECORD FOR AUSTRALIA. Muelleria 6(5):311-316 c6 -----1987b. NYMPHOIDES BELEGENSIS. Mulleria 6(5): 359-362 c4 -----1997. NYMPHOIDES SPINULOSPERMA: A NEW SPECIES FROM S.E. -

The Strength of Negative Plant–Soil Feedback Increases from the Intraspecific to the Interspecific and the Functional Group Level
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Institutional Repository of the Freie Universität Berlin Received: 14 July 2017 | Revised: 20 November 2017 | Accepted: 21 November 2017 DOI: 10.1002/ece3.3755 ORIGINAL RESEARCH The strength of negative plant–soil feedback increases from the intraspecific to the interspecific and the functional group level Alexandra R. Bukowski1 | Conrad Schittko2,3 | Jana S. Petermann3,4 1Institute of Biology, Freie Universität Berlin, Berlin, Germany Abstract 2Institute of Biochemistry and One of the processes that may play a key role in plant species coexistence and ecosys- Biology, Biodiversity Research/Systematic tem functioning is plant–soil feedback, the effect of plants on associated soil commu- Botany, University of Potsdam, Potsdam, Germany nities and the resulting feedback on plant performance. Plant–soil feedback at the 3Berlin-Brandenburg Institute of Advanced interspecific level (comparing growth on own soil with growth on soil from different Biodiversity Research (BBIB), Berlin, Germany species) has been studied extensively, while plant–soil feedback at the intraspecific 4Department of Ecology and Evolution, University of Salzburg, Salzburg, level (comparing growth on own soil with growth on soil from different accessions Austria within a species) has only recently gained attention. Very few studies have investi- Correspondence gated the direction and strength of feedback among different taxonomic levels, and Jana S. Petermann, Berlin-Brandenburg -

Thermogenesis in Three Philodendron Species (Araceae) of French Guiana Marc Gibernau1 and Denis Barabé2
Thermogenesis in three Philodendron species (Araceae) of French Guiana Marc Gibernau1 and Denis Barabé2 1. Laboratoire d’Ecologie Terrestre, Université Paul Sabatier, 118 Route de Narbonne, Bat 4R3, 31062 Toulosue cedex 4, France. E-mail: [email protected] 2. Institut de Recherche en Biologie Végétale, Université de Montréal, Jardin Botanique de Montréal, 4101 rue Sherbrooke Est, Montréal (Québec), Canada H1X 2B2. Abstract Spadix temperature was measured in three species of Philodendron: P. acutatum, P. pedatum and P. solimoesense. These species showed two different patterns of spadix temperature during their flowering cycle. In P. acutatum and P. pedatum (subgenus Philodendron), the spadix warmed up twice during the beginning of each flowering night with a temperature not significantly different from that of ambient air between the two peaks. In P. solimoesense (subgenus Meconostigma), the spadix temperature rose up to 14oC above that of ambient air during the first night, then it progressively cooled down but remained 3-6oC above ambient air temperature. We propose that the heat production and the spadix temperature patterns observed may reflect different physiological processes and have a taxonomic significance in the genus Philodendron. Keywords: Araceae, flowering cycle, flower temperature, heating flower. Résumé Nous avons mesuré la température du spadice chez trois espèces de Philodendron: P. acutatum, P. pedatum et P. solimoesense. Deux types de courbe de température des spadices ont été observés. Les spadices de P. acutatum, P. pedatum (sous-genre Philodendron) produisent deux pics distincts de chaleur lors des deux soirs de la floraison. Entre ces pics de chaleur, la température du spadice n’est pas différente de celle de l’air ambient. -
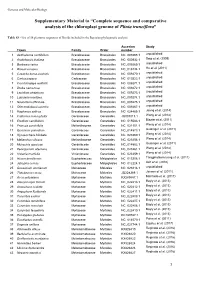
Complete Sequence and Comparative Analysis of the Chloroplast Genome of Plinia Trunciflora”
Genetics and Molecular Biology Supplementary Material to “Complete sequence and comparative analysis of the chloroplast genome of Plinia trunciflora” Table S3 - List of 56 plastome sequences of Rosids included in the Bayesian phylogenetic analysis. Accesion Study Taxon Family Order number 1 Aethionema cordifolium Brassicaceae Brassicales NC_009265.1 unpublished 2 Arabidopsis thaliana Brassicaceae Brassicales NC_000932.1 Sato et al. (1999) 3 Barbarea verna Brassicaceae Brassicales NC_009269.1 unpublished 4 Brassica napus Brassicaceae Brassicales NC_016734.1 Hu et al. (2011) 5 Capsella bursa-pastoris Brassicaceae Brassicales NC_009270.1 unpublished 6 Carica papaya Caricaceae Brassicales NC_010323.1 unpublished 7 Crucihimalaya wallichii Brassicaceae Brassicales NC_009271.1 unpublished 8 Draba nemorosa Brassicaceae Brassicales NC_009272.1 unpublished 9 Lepidium virginicum Brassicaceae Brassicales NC_009273.1 unpublished 10 Lobularia maritima Brassicaceae Brassicales NC_009274.1 unpublished 11 Nasturtium officinale Brassicaceae Brassicales NC_009275.1 unpublished 12 Olimarabidopsis pumila Brassicaceae Brassicales NC_009267.1 unpublished 13 Raphanus sativus Brassicaceae Brassicales NC_024469.1 Jeong et al. (2014) 14 California macrophylla Geraniaceae Geraniales JQ031013.1 Weng et al. (2014) 15 Erodium carvifolium Geraniaceae Geraniales NC_015083.1 Blazier et al. (2011) 16 Francoa sonchifolia Melianthaceae Geraniales NC_021101.1 Weng et al. (2014) 17 Geranium palmatum Geraniaceae Geraniales NC_014573.1 Guisinger et al. (2011) 18 Hypseocharis bilobate Geraniaceae Geraniales NC_023260.1 Weng et al. (2014) 19 Melianthus villosus Melianthaceae Geraniales NC_023256.1 Weng et al. (2014) 20 Monsonia speciose Geraniaceae Geraniales NC_014582.1 Guisinger et al. (2011) 21 Pelargonium alternans Geraniaceae Geraniales NC_023261.1 Weng et al. (2014) 22 Viviania marifolia Vivianiaceae Geraniales NC_023259.1 Weng et al. (2014) 23 Hevea brasiliensis Euphorbiaceae Malpighiales NC_015308.1 Tangphatsornruang et al. -

Downloaded from Genbank on That Full Plastid Genomes Are Not Sufficient to Reject Al- February 28, 2012
Ruhfel et al. BMC Evolutionary Biology 2014, 14:23 http://www.biomedcentral.com/1471-2148/14/23 RESEARCH ARTICLE Open Access From algae to angiosperms–inferring the phylogeny of green plants (Viridiplantae) from 360 plastid genomes Brad R Ruhfel1*, Matthew A Gitzendanner2,3,4, Pamela S Soltis3,4, Douglas E Soltis2,3,4 and J Gordon Burleigh2,4 Abstract Background: Next-generation sequencing has provided a wealth of plastid genome sequence data from an increasingly diverse set of green plants (Viridiplantae). Although these data have helped resolve the phylogeny of numerous clades (e.g., green algae, angiosperms, and gymnosperms), their utility for inferring relationships across all green plants is uncertain. Viridiplantae originated 700-1500 million years ago and may comprise as many as 500,000 species. This clade represents a major source of photosynthetic carbon and contains an immense diversity of life forms, including some of the smallest and largest eukaryotes. Here we explore the limits and challenges of inferring a comprehensive green plant phylogeny from available complete or nearly complete plastid genome sequence data. Results: We assembled protein-coding sequence data for 78 genes from 360 diverse green plant taxa with complete or nearly complete plastid genome sequences available from GenBank. Phylogenetic analyses of the plastid data recovered well-supported backbone relationships and strong support for relationships that were not observed in previous analyses of major subclades within Viridiplantae. However, there also is evidence of systematic error in some analyses. In several instances we obtained strongly supported but conflicting topologies from analyses of nucleotides versus amino acid characters, and the considerable variation in GC content among lineages and within single genomes affected the phylogenetic placement of several taxa. -

An Example from the Floral Epidermis Ofthe Nymphaeaceae
Zurich Open Repository and Archive University of Zurich Main Library Strickhofstrasse 39 CH-8057 Zurich www.zora.uzh.ch Year: 2018 Disentangling historical signal and pollinator selection on the micromorphology of flowers: an example from the floral epidermis ofthe Nymphaeaceae Coiro, Mario ; Barone Lumaga, Maria Rosaria Abstract: The family Nymphaeaceae includes most of the diversity among the ANA‐grade angiosperms. Among the species of this family, floral structures and pollination strategies are quite varied. The genus Victoria, as well as subgenera Lotos and Hydrocallis in Nymphaea, presents night‐blooming, scented flowers pollinated by scarab beetles. Such similar pollination strategies have led to macromorphological similarities among the flowers of these species, which could be interpreted as homologies or convergences based on different phylogenetic hypotheses about the relationships of these groups. We employed SEM of floral epidermis for seven species of the Nymphaeaceae with contrasting pollination biology to identify the main characters of the floral organs and the potential homologous nature of the structures involved in pollinator attraction. Moreover, we used TEM to observe ultrastructure of papillate‐conical epidermis in the stamen of Victoria cruziana. We then tested the phylogenetic or ecological distribution of these traits using both consensus network approaches and ancestral state reconstruction on fixed phylogenies. Our results show that the night‐blooming flowers present different specializations in their epidermis, with Victoria cruziana presenting the most elaborate floral anatomy. We also identify for the first timethe presence of conical‐papillate cells in the order Nymphaeales. The epidermal characters tend to reflect phylogenetic relationships more than convergence due to pollinator selection.