The Vascular Effects of Isolated Isoflavones—A Focus on The
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Protection of PC12 Cells Against Superoxide-Induced Damage by Isoflavonoids from Astragalus Mongholicus1
BIOMEDICAL AND ENVIRONMENTAL SCIENCES 22, 50-54 (2009) www.besjournal.com Protection of PC12 Cells against Superoxide-induced Damage by 1 Isoflavonoids from Astragalus mongholicus # + + #,2 DE-HONG YU , YONG-MING BAO , LI-JIA AN , AND MING YANG #The State Key Laboratory of Natural and Biomimetic Drugs, Peking University, Beijing 100083, China; +Department of Bioscience and Biotechnology, Dalian University of Technology, Dalian 116024, Liaoning, China Objective To further investigate the neuroprotective effects of five isoflavonoids from Astragalus mongholicus on xanthine (XA)/ xanthine oxidase (XO)-induced injury to PC12 cells. Methods PC12 cells were damaged by XA/XO. The activities of antioxidant enzymes, MTT, LDH, and GSH assays were used to evaluate the protection of these five isoflavonoids. Contents of Bcl-2 family proteins were determined with flow cytometry. Results Among the five isoflavonoids including formononetin, ononin, 9, 10-dimethoxypterocarpan-3-O-β-D-glucoside, calycosin and calycosin-7-O-glucoside, calycosin and calycosin-7-O-glucoside were found to inhibit XA/ XO-induced injury to PC12 cells. Their EC50 values of formononetin and calycosin were 0.05 μg/mL. Moreover, treatment with these three isoflavonoids prevented a decrease in the activities of antioxidant enzymes, superoxide dismutase (SOD) and glutathione peroxidase (GSH-Px), while formononetin and calycosin could prevent a significant deletion of GSH. In addition, only calycosin and calycosin-7-O-glucoside were shown to inhibit XO activity in cell-free system, with an approximate IC50 value of 10 μg/mL and 50 μg/mL. Formononetin and calycosin had no significant influence on Bcl-2 or Bax protein contents. Conclusion Neuroprotection of formononetin, calycosin and calycosin-7-O-glucoside may be mediated by increasing endogenous antioxidants, rather by inhibiting XO activities or by scavenging free radicals. -

Content and Composition of Isoflavonoids in Mature Or Immature
394 Journal of Health Science, 47(4) 394–406 (2001) Content and Composition of tivities1) and reported to be protective against can- cer, cardiovascular diseases and osteoporosis.3–9) Isoflavonoids in Mature or Much research has been reported about the content Immature Beans and Bean of isoflavonoids in soybeans and soybean-derived processed foods.10–23) In contrast, there are few re- Sprouts Consumed in Japan ports about the isoflavonoid content in beans other than soybeans.11,12,18,23) Yumiko Nakamura,* Akiko Kaihara, Japanese people are reported to ingest Kimihiko Yoshii, Yukari Tsumura, isoflavonoids mainly through the consumption of Susumu Ishimitsu, and Yasuhide Tonogai soybeans and its derived processed foods.20) Re- cently, we estimated that the Japanese daily intake Division of Food Chemistry, National Institute of Health Sci- of isoflavonoids from soybeans and soybean-based ences, Osaka Branch, 1–1–43 Hoenzaka, Chuo-ku, Osaka 540– processed foods is 27.80 mg per day (daidzein 0006, Japan (Received January 9, 2001; Accepted April 6, 2001) 12.02 mg, glycitein 2.30 mg and genistein 13.48 mg).24) However, isoflavonoid intake from the The content of 9 types of isoflavonoids (daidzein, consumption of immature beans, sprouts and beans glycitein, genistein, formononetin, biochanin A, other than soybeans has not been elucidated. Here coumestrol, daidzin, glycitin and genistin) in 34 do- we have measured the content of isoflavonoids in mestic or imported raw beans including soybeans, 7 mature and immature beans and bean sprouts con- immature beans and 5 bean sprouts consumed in Ja- sumed in Japan, and have compared the content and pan were systematically analyzed. -

Interpreting Sources and Endocrine Active Components of Trace Organic Contaminant Mixtures in Minnesota Lakes
INTERPRETING SOURCES AND ENDOCRINE ACTIVE COMPONENTS OF TRACE ORGANIC CONTAMINANT MIXTURES IN MINNESOTA LAKES by Meaghan E. Guyader © Copyright by Meaghan E. Guyader, 2018 All Rights Reserved A thesis submitted to the Faculty and the Board of Trustees of the Colorado School of Mines in partial fulfillment of the requirements for the degree of Doctor of Philosophy (Civil and Environmental Engineering). Golden, Colorado Date _____________________________ Signed: _____________________________ Meaghan E. Guyader Signed: _____________________________ Dr. Christopher P. Higgins Thesis Advisor Golden, Colorado Date _____________________________ Signed: _____________________________ Dr. Terri S. Hogue Professor and Department Head Department of Civil and Environmental Engineering ii ABSTRACT On-site wastewater treatment systems (OWTSs) are a suspected source of widespread trace organic contaminant (TOrC) occurrence in Minnesota lakes. TOrCs are a diverse set of synthetic and natural chemicals regularly used as cleaning agents, personal care products, medicinal substances, herbicides and pesticides, and foods or flavorings. Wastewater streams are known to concentrate TOrC discharges to the environment, particularly accumulating these chemicals at outfalls from centralized wastewater treatment plants. Fish inhabiting these effluent dominated environments are also known to display intersex qualities. Concurrent evidence of this phenomenon, known as endocrine disruption, in Minnesota lake fish drives hypotheses that OWTSs, the primary form of wastewater treatment in shoreline residences, may contribute to TOrC occurrence and the endocrine activity in these water bodies. The causative agents specific to fish in this region remain poorly understood. The objective of this dissertation was to investigate OWTSs as sources of TOrCs in Minnesota lakes, and TOrCs as potential causative agents for endocrine disruption in resident fish. -

Suspect and Target Screening of Natural Toxins in the Ter River Catchment Area in NE Spain and Prioritisation by Their Toxicity
toxins Article Suspect and Target Screening of Natural Toxins in the Ter River Catchment Area in NE Spain and Prioritisation by Their Toxicity Massimo Picardo 1 , Oscar Núñez 2,3 and Marinella Farré 1,* 1 Department of Environmental Chemistry, IDAEA-CSIC, 08034 Barcelona, Spain; [email protected] 2 Department of Chemical Engineering and Analytical Chemistry, University of Barcelona, 08034 Barcelona, Spain; [email protected] 3 Serra Húnter Professor, Generalitat de Catalunya, 08034 Barcelona, Spain * Correspondence: [email protected] Received: 5 October 2020; Accepted: 26 November 2020; Published: 28 November 2020 Abstract: This study presents the application of a suspect screening approach to screen a wide range of natural toxins, including mycotoxins, bacterial toxins, and plant toxins, in surface waters. The method is based on a generic solid-phase extraction procedure, using three sorbent phases in two cartridges that are connected in series, hence covering a wide range of polarities, followed by liquid chromatography coupled to high-resolution mass spectrometry. The acquisition was performed in the full-scan and data-dependent modes while working under positive and negative ionisation conditions. This method was applied in order to assess the natural toxins in the Ter River water reservoirs, which are used to produce drinking water for Barcelona city (Spain). The study was carried out during a period of seven months, covering the expected prior, during, and post-peak blooming periods of the natural toxins. Fifty-three (53) compounds were tentatively identified, and nine of these were confirmed and quantified. Phytotoxins were identified as the most frequent group of natural toxins in the water, particularly the alkaloids group. -

1 1 2 3 4 5 6 7 Bioactive Phenolic Compounds of Soybean
* Manuscript Click here to view linked References 1 Bioactive phenolic compounds of soybean ( Glycine max cv. Merit): modifications by 2 different microbial fermentations 3 4 5 M. Dueñas 2; T. Hernández 1; S. Robredo 1; *I. Estrella 1; R. Muñoz 1 6 7 1Instituto de Fermentaciones Industriales. C.S.I.C. Juan de la Cierva 3, 28006-Madrid. Spain. Fax: 34-915644853. 8 [email protected] 9 2Grupo de Investigación en Polifenoles, Unidad de Nutrición y Bromatología, Facultad de Farmacia, Universidad de 10 Salamanca. Campus Miguel de Unamuno. 37007 Salamanca. Spain. 11 12 *Corresponding author 13 14 Running title: Changes in phenolic compounds of soybeans by microbial fermentations 15 16 17 Abstract 18 Phenolic compounds of soybeans ( Glycine max ) are partially responsible of their beneficial 19 health effects. The most abundant phenolics in soybeans are isoflavones, mainly daidzein, 20 glycitein, and genistein and several derivatives of them. Other phenolic compounds are also 21 present but they have been minimally studied. In this work, the soybean seeds were fermented by 22 the microbiota present on the seeds or by inoculation with Aspergillus oryzae, Rhizopus oryzae or 23 Bacillus subtilis . The effect of microbial fermentation on the phenolic composition of soybeans 24 was studied. All the assayed microorganisms brought out variations in phenolics. An increase in 25 the concentration of phenolic acids was observed. Flavonoids were differently metabolized 26 depending on the microorganism used. In all samples the most important changes were produced 27 in the isoflavones content, observing an increase on the aglycones, glycitein, daidzein and 28 genistein. 29 30 Key words : soybean, fermentation, phenolic compounds, isoflavones 31 1 32 1. -
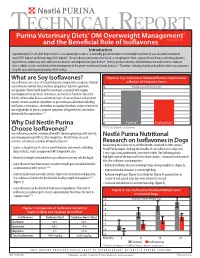
What Are Soy Isoflavones? Figure 2: Soy Isoflavones Reduced Plasma Isoprostanes Soy Isoflavones Are a Class of Natural Bioactive Compounds in Soybeans
TECHNICAL REPORT Purina Veterinary Diets® OM Overweight Management® brand canine dry formula and the Beneficial Role of Isoflavones Introduction Approximately 35% of adult dogs in the U.S. are overweight or obese1. A markedly greater incidence of overweight and obesity was observed in neutered male (59% higher) and female dogs (40% higher)1. Chronic obesity can increase the risk of, or complications from, several chronic diseases including diabetes, hypertension, pulmonary and cardiovascular disease, and degenerative joint disease2. Obesity produces chronic, mild inflammation and increases oxidative stress, which, in turn, contributes to the development of the above-mentioned chronic diseases.3,4 Therefore, reducing obesity and oxidative stress can promote a long life span and improved quality of life in dogs. What are Soy Isoflavones? Figure 2: Soy Isoflavones Reduced Plasma Isoprostanes Soy isoflavones are a class of natural bioactive compounds in soybeans. Natural a Marker of Oxidative Stress soy isoflavones include three chemical compounds: daidzein, genistein, 5 Plasma Isoprostanes (ng/ml) and glycitein. Many health benefits have been associated with regular consumption of soy products. In humans, soy has been found to reduce the 4 risk of cardiovascular disease and certain types of cancer (breast and prostate cancer); relieve a number of problems in post menopausal women including 3 hot flashes, osteoporosis, and decline in cognitive function; reduce cholesterol and triglycerides in plasma; improve symptoms of hypertension; and reduce 2 abdominal fat accumulation.5,6,7 1 Why Did Nestlé Purina 0 Control Isoflavones* Choose Isoflavones? *P<0.01 for Control vs. Isoflavones Soy isoflavones provide a number of benefits for managing dogs with obesity, and reducing rebound effects after weight loss. -

Plant Phenolics: Bioavailability As a Key Determinant of Their Potential Health-Promoting Applications
antioxidants Review Plant Phenolics: Bioavailability as a Key Determinant of Their Potential Health-Promoting Applications Patricia Cosme , Ana B. Rodríguez, Javier Espino * and María Garrido * Neuroimmunophysiology and Chrononutrition Research Group, Department of Physiology, Faculty of Science, University of Extremadura, 06006 Badajoz, Spain; [email protected] (P.C.); [email protected] (A.B.R.) * Correspondence: [email protected] (J.E.); [email protected] (M.G.); Tel.: +34-92-428-9796 (J.E. & M.G.) Received: 22 October 2020; Accepted: 7 December 2020; Published: 12 December 2020 Abstract: Phenolic compounds are secondary metabolites widely spread throughout the plant kingdom that can be categorized as flavonoids and non-flavonoids. Interest in phenolic compounds has dramatically increased during the last decade due to their biological effects and promising therapeutic applications. In this review, we discuss the importance of phenolic compounds’ bioavailability to accomplish their physiological functions, and highlight main factors affecting such parameter throughout metabolism of phenolics, from absorption to excretion. Besides, we give an updated overview of the health benefits of phenolic compounds, which are mainly linked to both their direct (e.g., free-radical scavenging ability) and indirect (e.g., by stimulating activity of antioxidant enzymes) antioxidant properties. Such antioxidant actions reportedly help them to prevent chronic and oxidative stress-related disorders such as cancer, cardiovascular and neurodegenerative diseases, among others. Last, we comment on development of cutting-edge delivery systems intended to improve bioavailability and enhance stability of phenolic compounds in the human body. Keywords: antioxidant activity; bioavailability; flavonoids; health benefits; phenolic compounds 1. Introduction Phenolic compounds are secondary metabolites widely spread throughout the plant kingdom with around 8000 different phenolic structures [1]. -

Antiproliferative Effects of Isoflavones on Human Cancer Cell Lines Established from the Gastrointestinal Tract 1
[CANCER RESEARCH 53, 5815-5821, December 1, 1993] Antiproliferative Effects of Isoflavones on Human Cancer Cell Lines Established from the Gastrointestinal Tract 1 Kazuyoshi Yanagihara, 2 Akihiro Ito, Tetsuya Toge, and Michitaka Numoto Department of Pathology [K. Y, M. N.], Cancer Research [A. L], and Surgery iT. T.], Research Institute for Nuclear Medicine and Biology, Hiroshima University, Kasumi 1-2-3, Minami-ku, Hiroshima 734, Japan ABSTRACT MATERIALS AND METHODS Seven isoflavones, biochanin A, daidzein, genistein, genistin, prunectin, Establishment of Cancer Cell Lines. Tumor tissues were trimmed of fat puerarin, and pseudobaptigenin were tested for cytostatic and cytotoxic and necrotic portions and minced with scalpels. The tissue pieces were trans- effects on 10 newly established cancer cell lines of the human gastrointes- ferred, together with medium, at 10 to 15 fragments/dish to 60-mm culture tinal origin. Proliferation of HSC-41E6, HSC-45M2, and SH101-P4 stom- dishes (Falcon, Lincoln Park, N J). The patient's ascitic tumor cells were ach cancer cell lines was strongly inhibited by biochanin A and genistein, harvested by centrifugation (1000 rpm for 10 min) and plated into 60-mm whereas other stomach, esophageal, and colon cancer lines were moder- dishes. Culture dishes initially selectively trypsinized [trypsin, 0.05% (w/v); ately suppressed by both compounds. Biochanin A and genistein were EDTA, 0.02% (w/v)], to remove overgrowing fibroblasts. In addition, we cytostatic at low concentrations (<20 pg/ml for biochanin A, <10/tg/ml for attempted to remove the fibroblasts mechanically and to transfer the tumor genistein) and were cytotoxic at higher concentrations (>40 pg/ml for cells selectively. -

Applications of in Silico Methods to Analyze the Toxicity and Estrogen T Receptor-Mediated Properties of Plant-Derived Phytochemicals ∗ K
Food and Chemical Toxicology 125 (2019) 361–369 Contents lists available at ScienceDirect Food and Chemical Toxicology journal homepage: www.elsevier.com/locate/foodchemtox Applications of in silico methods to analyze the toxicity and estrogen T receptor-mediated properties of plant-derived phytochemicals ∗ K. Kranthi Kumara, P. Yugandharb, B. Uma Devia, T. Siva Kumara, N. Savithrammab, P. Neerajaa, a Department of Zoology, Sri Venkateswara University, Tirupati, 517502, India b Department of Botany, Sri Venkateswara University, Tirupati, 517502, India ARTICLE INFO ABSTRACT Keywords: A myriad of phytochemicals may have potential to lead toxicity and endocrine disruption effects by interfering Phytochemicals with nuclear hormone receptors. In this examination, the toxicity and estrogen receptor−binding abilities of a QSAR modeling set of 2826 phytochemicals were evaluated. The endpoints mutagenicity, carcinogenicity (both CAESAR and ISS Toxicity models), developmental toxicity, skin sensitization and estrogen receptor relative binding affinity (ER_RBA) Nuclear hormone receptor binding were studied using the VEGA QSAR modeling package. Alongside the predictions, models were providing pos- Self−Organizing maps sible information for applicability domains and most similar compounds as similarity sets from their training Clustering and classification schemes sets. This information was subjected to perform the clustering and classification of chemicals using Self−Organizing Maps. The identified clusters and their respective indicators were considered as potential hotspot structures for the specified data set analysis. Molecular screening interpretations of models wereex- hibited accurate predictions. Moreover, the indication sets were defined significant clusters and cluster in- dicators with probable prediction labels (precision). Accordingly, developed QSAR models showed good pre- dictive abilities and robustness, which observed from applicability domains, representation spaces, clustering and classification schemes. -

Insect-Induced Daidzein, Formononetin and Their Conjugates in Soybean Leaves
UC San Diego UC San Diego Previously Published Works Title Insect-induced daidzein, formononetin and their conjugates in soybean leaves. Permalink https://escholarship.org/uc/item/5pw0t3dx Journal Metabolites, 4(3) ISSN 2218-1989 Authors Murakami, Shinichiro Nakata, Ryu Aboshi, Takako et al. Publication Date 2014-07-04 DOI 10.3390/metabo4030532 Peer reviewed eScholarship.org Powered by the California Digital Library University of California Metabolites 2014, 4, 532-546; doi:10.3390/metabo4030532 OPEN ACCESS metabolites ISSN 2218-1989 www.mdpi.com/journal/metabolites/ Article Insect-Induced Daidzein, Formononetin and Their Conjugates in Soybean Leaves Shinichiro Murakami 1, Ryu Nakata 1, Takako Aboshi 1, Naoko Yoshinaga 1, Masayoshi Teraishi 1, Yutaka Okumoto 1, Atsushi Ishihara 3, Hironobu Morisaka 1, Alisa Huffaker 2, Eric A Schmelz 2 and Naoki Mori 1,* 1 Graduate School of Agriculture, Kyoto University, Kitashirakawa, Sakyo, Kyoto 606-8502, Japan; E-Mails: [email protected] (S.M.); [email protected] (R.N.); [email protected] (T.A.); [email protected] (N.Y.); [email protected] (M.T.); [email protected] (Y.O.); [email protected] (H.M.) 2 Center for Medical, Agricultural, and Veterinary Entomology, Agricultural Research Service, USDA, 1600 S.W. 23RD Drive, Gainesville, FL 32606, USA; E-Mails: [email protected] (A.H.); [email protected] (E.A.S.) 3 Department of Agriculture, Tottori University, Koyama-machi 4-101, Tottori 680-8550, Japan; E-Mail: [email protected] * Author to whom correspondence should be addressed; E-Mail: [email protected]; Tel.: +81-75-753-6307. -

Daidzein and Genistein Content of Cereals
European Journal of Clinical Nutrition (2002) 56, 961–966 ß 2002 Nature Publishing Group All rights reserved 0954–3007/02 $25.00 www.nature.com/ejcn ORIGINAL COMMUNICATION Daidzein and genistein content of cereals J Liggins1, A Mulligan1,2, S Runswick1 and SA Bingham1,2* 1Medical Research Council Dunn Human Nutrition Unit, Hills Road, Cambridge, UK; and 2European Prospective Investigation of Cancer, University of Cambridge, Cambridge, UK Objective: To analyse 75 cereals and three soy flours commonly eaten in Europe for the phytoestrogens daidzein and genistein. Design: The phytoestrogens daidzein and genistein were extracted from dried foods, and the two isoflavones quantified after hydrolytic removal of any conjugated carbohydrate. Completeness of extraction and any procedural losses of the isoflavones 0 0 were accounted for using synthetic daidzin (7-O-glucosyl-4 -hydroxyisoflavone) and genistin (7-O-glucosyl-4 5-dihydroxyiso- flavone) as internal standards. Setting: Foods from the Cambridge UK area were purchased, prepared for eating, which included cooking if necessary, and freeze dried. Three stock soy flours were also analysed. Results: Eighteen of the foods assayed contained trace or no detectable daidzein or genistein. The soy flours were rich sources, containing 1639 – 2117 mg=kg. The concentration of the two isoflavones in the remaining foods ranged from 33 to 11 873 mg=kg. Conclusion: These analyses will supply useful information to investigators determining the intake of phytoestrogens in cereal products in order to relate intakes to potential biological activities. Sponsorship: This work was supported by the United Kingdom Medical Research Council, Ministry of Agriculture Fisheries and Food (contract FS2034) and the United States of America Army (contract DAMD 17-97-1-7028). -

Mindy Goldman, MD Clinical Professor Dept
Managing Menopause Medically and Naturally Mindy Goldman, MD Clinical Professor Dept. of Ob/Gyn and Reproductive Sciences Director, Women’s Cancer Care Program, UCSF Breast Care Center and Women’s Health University of California, San Francisco I have nothing to disclose –Mindy Goldman, MD CASE STUDY 50 yr. old G2P2 peri-menopausal woman presents with complaints of significant night sweats interfering with her ability to sleep. She has mild hot flashes during the day. She has never had a bone mineral density test but her mother had a hip fracture at age 62 due to osteoporosis. Her 46 yr. old sister was diagnosed with breast cancer at age 43, treated with lumpectomy and radiation and currently is doing well. There is no other family history of cancer. Questions 1. Would you offer her MHT? 2. If yes, how long would you continue it? 3. If no, what would you offer for alternative treatments? 4. Would your treatment differ if you knew she had underlying heart disease? Is it safe? How long can I take it? What about Mymy Bones?bones? Will it protect my heart? MHT - 2015 What about my brain? Will I get breast cancer? What about my hot flashes? Menopausal Symptoms Hot flashes Night sweats Sleep disturbances Vaginal dryness/Sexual dysfunction Mood disturbances How to Treat Menopausal Symptoms Hormone therapy Alternatives to hormones Complementary and Integrative Techniques Prior to Women’s Health Initiative Hormone therapy primary treatment of menopausal hot flashes Few women would continue hormones past one year By 1990’s well known