Pdf) on July 4, 2021 [Editor: Bruno R.C
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Xenosphinctes N. Gen. (Ataxioceratidae, Lithacoceratinae), a New Rare Ammonite Genus from the Lower Tithonian (Hybonotum Zone) of SW Germany
VOLUMINA JURASSICA, 2017, XV: 155–160 DOI: 10.5604/01.3001.0010.7170 Xenosphinctes n. gen. (Ataxioceratidae, Lithacoceratinae), a new rare ammonite genus from the Lower Tithonian (Hybonotum Zone) of SW Germany Armin SCHERZINGER1, Günter SCHWEIGERT2 Key words: ammonites, Xenosphinctes berkai, Early Tithonian, biostratigraphy, Germany. Abstract. The new monotypic ammonite genus Xenosphinctes (type species: Xenosphinctes berkai n. sp.) is established. It is recorded from the Upper Jurassic, Early Tithonian, Hybonotum Zone, Riedense Subzone, eigeltingense α horizon from the Talmühle, N of Engen, Baden-Württemberg, SW Germany. INTRODUCTION AMMONITE FAUNA OF THE EIGELTINGENSE α HORIZON The Hangende-Bankkalke Formation is the youngest formation in the Upper Jurassic of Swabia. Its base is usual- The first comprehensive review of ammonites from the ly formed by an unconformity with a marked change in li- uppermost Jurassic in Swabia was provided by Berckhemer thology from coarse-grained bioclastic limestones or marl- & Hölder (1959). However, therein the focus was on the am- stones to pure mudstones (Schweigert, 1996). Its type monite fauna of the Late Kimmeridgian Beckeri Zone, and locality is an abandoned limestone quarry near the village of only very few taxa from higher levels have been reported. Eigeltingen, and its base is best exposed in a nearby section Ohmert & Zeiss (1980) started with a description of the am- along a railroad cut, NW of the Talmühle hamlet (Fig. 1). monite fauna of the Hangende-Bankkalke Formation in the The most abundant macrofossils in these basal parts of the western part of the Swabian Alb. On that occasion several formation are ammonites. Many of them are fragmented due important taxa from the basal part of this formation which to predation, but some of them are quite well-preserved as are typical of the former eigeltingense horizon were intro- steinkerns with superimposed sculpture of the outer shell. -

First Three-Dimensionally Preserved in Situ Record of an Aptychophoran Ammonite Jaw Apparatus in the Jurassic and Discussion of the Function of Aptychi
Berliner paläobiologische Abhandlungen 10 321-330 Berlin 2009-11-11 First three-dimensionally preserved in situ record of an aptychophoran ammonite jaw apparatus in the Jurassic and discussion of the function of aptychi Günter Schweigert Abstract: A unique specimen of the microconch ammonite Lingulaticeras planulatum Berckhemer in Ziegler, 1958 comes from a tempestite bed within the Upper Jurassic lithographic limestones of Scham- haupten in Franconia (Painten Formation, uppermost Kimmeridgian). The shell is unique because it retains the complete jaw apparatus in the body chamber. The articulation of the Lamellaptychus and the corresponding upper beak are well preserved. The function of the aptychus is discussed in general, and an operculum function is thought to be unlikely. The formation of strongly calcified aptychi in aspidoceratids and some oppeliid ammonoids is interpreted as an added ballast weight to stabilize the conch for swimming in the water column. Keywords: Ammonites, aptychus, preservation, functional morphology, Upper Jurassic, lithographic lime- stones, Franconia, Germany Zusammenfassung: Ein einzigartig erhaltenes Exemplar des mikroconchen Ammoniten Lingulaticeras planulatum Berckhemer in Ziegler, 1958 aus einer Tempestitlage des oberjurassischen Plattenkalks von Schamhaupten in Franken (Painten-Formation, oberstes Kimmeridgium) enthält noch den vollständigen Kieferapparat in seiner Wohnkammer.Es zeigt die perfekte Artikualation des Lamellaptychus mit dem dazu- gehörenden Oberkiefer. Die Funktion des Aptychus wird allgemein diskutiert und eine Deckelfunktion für unwahrscheinlich gehalten. Die Ausbildung stark verkalkter Aptychen wie in Aspidoceraten und manchen Oppeliiden wird als zusätzliches Tariergewicht gedeutet, um das Gehäuse in starker bewegtem Wasser zu stabilisieren. Schlüsselwörter: Ammoniten, Aptychus, Erhaltung, Plattenkalke, Funktionsmorphologie, Oberjura, Franken, Deutschland Address of the author: Dr. Günter Schweigert, Staatliches Museum für Naturkunde, Rosenstein 1, D-70191 Stuttgart. -
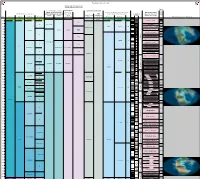
Expanded Jurassic Timescale
TimeScale Creator 2012 chart Russian and Ural regional units Russia Platform regional units Calca Jur-Cret boundary regional Russia Platform East Asian regional units reous stages - British and Boreal Stages (Jur- Australia and New Zealand regional units Marine Macrofossils Nann Standard Chronostratigraphy British regional Boreal regional Cret, Perm- Japan New Zealand Chronostratigraphy Geomagnetic (Mesozoic-Paleozoic) ofossil stages stages Carb & South China (Neogene & Polarity Tethyan Ammonoids s Ma Period Epoch Age/Stage Substage Cambrian) stages Cret) NZ Series NZ Stages Global Reconstructions (R. Blakey) Ryazanian Ryazanian Ryazanian [ no stages M17 CC2 Cretaceous Early Berriasian E Kochian Taitai Um designated ] M18 CC1 145 Berriasella jacobi M19 NJT1 Late M20 7b 146 Lt Portlandian M21 Durangites NJT1 M22 7a 147 Oteke Puaroan Op M22A Micracanthoceras microcanthum NJT1 Penglaizhenian M23 6b 148 Micracanthoceras ponti / Volgian Volgian Middle M24 Burckhardticeras peroni NJT1 Tithonian M24A 6a 149 M24B Semiformiceras fallauxi NJT15 M25 b E 150 M25A Semiformiceras semiforme NJT1 5a Early M26 Semiformiceras darwini 151 lt-Oxf N M-Sequence Hybonoticeras hybonotum 152 Ohauan Ko lt-Oxf R Kimmeridgian Hybonoticeras beckeri 153 m- Lt Late Oxf N Aulacostephanus eudoxus 154 m- NJT14 Late Aspidoceras acanthicum Oxf R Kimmeridgian Kimmeridgian Kimmeridgian Crussoliceras divisum 155 155.431 Card- N Ataxioceras hypselocyclum 156 E Early e-Oxf Sutneria platynota R Idoceras planula Suiningian 157 Cal- Oxf N Epipeltoceras bimammatum 158 lt- Lt Callo -

A Review of the Classification of Jurassic Aspidoceratid Ammonites – the Superfamily Aspidoceratoidea
VOLUMINA JURASSICA, 2020, XVIII (1): 47–52 DOI: 10.7306/VJ.18.4 A review of the classification of Jurassic aspidoceratid ammonites – the Superfamily Aspidoceratoidea Horacio PARENT1, Günter SCHWEIGERT2, Armin SCHERZINGER3 Key words: Superfamily Aspidoceratoidea, Aspidoceratidae, Epipeltoceratinae emended, Peltoceratidae, Gregoryceratinae nov. subfam. Abstract. The aspidoceratid ammonites have been traditionally included in the superfamily Perisphinctoidea. However, the basis of this is unclear for they bear unique combinations of characters unknown in typical perisphinctoids: (1) the distinct laevaptychus, (2) stout shells with high growth rate of the whorl section area, (3) prominent ornamentation with tubercles, spines and strong growth lines running in parallel over strong ribs, (4) lack of constrictions, (5) short to very short bodychamber, and (6) sexual dimorphism characterized by minia- turized microconchs and small-sized macroconchs besides the larger ones, including changes of sex during ontogeny in many cases. Considering the uniqueness of these characters we propose herein to raise the family Aspidoceratidae to the rank of a superfamily Aspi- doceratoidea, ranging from the earliest Late Callovian to the Early Berriasian Jacobi Zone. The new superfamily includes two families, Aspidoceratidae (Aspidoceratinae, Euaspidoceratinae, Epipeltoceratinae and Hybonoticeratinae), and Peltoceratidae (Peltoceratinae and Gregoryceratinae nov. subfam.). The highly differentiated features of the aspidoceratoids indicate that their life-histories -

Constraints on the Timescale of Animal Evolutionary History
Palaeontologia Electronica palaeo-electronica.org Constraints on the timescale of animal evolutionary history Michael J. Benton, Philip C.J. Donoghue, Robert J. Asher, Matt Friedman, Thomas J. Near, and Jakob Vinther ABSTRACT Dating the tree of life is a core endeavor in evolutionary biology. Rates of evolution are fundamental to nearly every evolutionary model and process. Rates need dates. There is much debate on the most appropriate and reasonable ways in which to date the tree of life, and recent work has highlighted some confusions and complexities that can be avoided. Whether phylogenetic trees are dated after they have been estab- lished, or as part of the process of tree finding, practitioners need to know which cali- brations to use. We emphasize the importance of identifying crown (not stem) fossils, levels of confidence in their attribution to the crown, current chronostratigraphic preci- sion, the primacy of the host geological formation and asymmetric confidence intervals. Here we present calibrations for 88 key nodes across the phylogeny of animals, rang- ing from the root of Metazoa to the last common ancestor of Homo sapiens. Close attention to detail is constantly required: for example, the classic bird-mammal date (base of crown Amniota) has often been given as 310-315 Ma; the 2014 international time scale indicates a minimum age of 318 Ma. Michael J. Benton. School of Earth Sciences, University of Bristol, Bristol, BS8 1RJ, U.K. [email protected] Philip C.J. Donoghue. School of Earth Sciences, University of Bristol, Bristol, BS8 1RJ, U.K. [email protected] Robert J. -

301083243.Pdf
AMEGHINIANA ISSN 0002-7014 Revista de la Asociación Paleontológica Argentina Tomo XII Diciembre de 1975 N?4 THE INDO-PACIFIC AMMONITE MAY AITES IN THE OXFORDIAN OF THESOUTHERN ANDES By P. N. STIPANICIC', G. E. G. WESTERMANN 2 and A. C. RJCCARDI3 ABSTRACT: Oxfordian Iitho- and biostratigraphy of the Chilean and Argentine Andes is reviewed (P. N. Stipanicic). Within the Chacay Group, the Lower to basal Upper Oxfordian La Manga Formation, below, mostly detrital and biogenic, and the Upper Oxfordian Au- quilco Formation, above, mainly chemical, are distinguished. The La Manga Formation (with Gryphaea calceola lumachelle) is rich in ammonite faunas, particularly of thc upper Cordatum to lower Canaliculatum Zones. In Neuquén and Mendoza provinces of Argentina, the Pli- catilis Zone or Middle Oxfordian has yielded Perísphinctes spp., Euaspidoceras spp., Aspido- ceras spp., together with Mayaítes (Araucanites ) stípanícfcí, M. (A.) reyesi, and M. (A.) mulai, Westermann et Riccardi subgen. et spp. nov. The first find of Mayaitidae outside the Indo-Pacific province is discussed in light of _plate-tectonic theory. RESUMEN: La revisron Iito- y bioestratigráfica del Oxfordiano de los Andes de Argentina y Chile (P. N. Stipanicic) ha permitido reconocer dentro del Grupo Chacay: 1) abajo, la Formación La Manga, mayormente detrítica y biogénica, del Oxfordiano inferior-superior basa!, y 2) arriba, la Formación Auquilco, mayormente química, del Oxfordiano superior. La For- mación La Manga (con lumachelas de Gryphaea calceola) contiene abundante cantidad de amonitas, particularmente de las Zonas de Cordatum superior a Canaliculatum inferior. En las provincias de Mendoza y Neuquén,Argentina, la Zona de Plicatilis (Oxfordiano medio) contiene Perispbinctes spp., Euaspidoceras spp., Aspidoceras spp., conjuntamente con Mayaites (Araucanites) stipanicici, M. -

Early Ontogeny of Jurassic Bakevelliids and Their Bearing on Bivalve Evolution
Early ontogeny of Jurassic bakevelliids and their bearing on bivalve evolution NIKOLAUS MALCHUS Malchus, N. 2004. Early ontogeny of Jurassic bakevelliids and their bearing on bivalve evolution. Acta Palaeontologica Polonica 49 (1): 85–110. Larval and earliest postlarval shells of Jurassic Bakevelliidae are described for the first time and some complementary data are given concerning larval shells of oysters and pinnids. Two new larval shell characters, a posterodorsal outlet and shell septum are described. The outlet is homologous to the posterodorsal notch of oysters and posterodorsal ridge of arcoids. It probably reflects the presence of the soft anatomical character post−anal tuft, which, among Pteriomorphia, was only known from oysters. A shell septum was so far only known from Cassianellidae, Lithiotidae, and the bakevelliid Kobayashites. A review of early ontogenetic shell characters strongly suggests a basal dichotomy within the Pterio− morphia separating taxa with opisthogyrate larval shells, such as most (or all?) Praecardioida, Pinnoida, Pterioida (Bakevelliidae, Cassianellidae, all living Pterioidea), and Ostreoida from all other groups. The Pinnidae appear to be closely related to the Pterioida, and the Bakevelliidae belong to the stem line of the Cassianellidae, Lithiotidae, Pterioidea, and Ostreoidea. The latter two superfamilies comprise a well constrained clade. These interpretations are con− sistent with recent phylogenetic hypotheses based on palaeontological and genetic (18S and 28S mtDNA) data. A more detailed phylogeny is hampered by the fact that many larval shell characters are rather ancient plesiomorphies. Key words: Bivalvia, Pteriomorphia, Bakevelliidae, larval shell, ontogeny, phylogeny. Nikolaus Malchus [[email protected]], Departamento de Geologia/Unitat Paleontologia, Universitat Autòno− ma Barcelona, 08193 Bellaterra (Cerdanyola del Vallès), Spain. -

Contributions in BIOLOGY and GEOLOGY
MILWAUKEE PUBLIC MUSEUM Contributions In BIOLOGY and GEOLOGY Number 51 November 29, 1982 A Compendium of Fossil Marine Families J. John Sepkoski, Jr. MILWAUKEE PUBLIC MUSEUM Contributions in BIOLOGY and GEOLOGY Number 51 November 29, 1982 A COMPENDIUM OF FOSSIL MARINE FAMILIES J. JOHN SEPKOSKI, JR. Department of the Geophysical Sciences University of Chicago REVIEWERS FOR THIS PUBLICATION: Robert Gernant, University of Wisconsin-Milwaukee David M. Raup, Field Museum of Natural History Frederick R. Schram, San Diego Natural History Museum Peter M. Sheehan, Milwaukee Public Museum ISBN 0-893260-081-9 Milwaukee Public Museum Press Published by the Order of the Board of Trustees CONTENTS Abstract ---- ---------- -- - ----------------------- 2 Introduction -- --- -- ------ - - - ------- - ----------- - - - 2 Compendium ----------------------------- -- ------ 6 Protozoa ----- - ------- - - - -- -- - -------- - ------ - 6 Porifera------------- --- ---------------------- 9 Archaeocyatha -- - ------ - ------ - - -- ---------- - - - - 14 Coelenterata -- - -- --- -- - - -- - - - - -- - -- - -- - - -- -- - -- 17 Platyhelminthes - - -- - - - -- - - -- - -- - -- - -- -- --- - - - - - - 24 Rhynchocoela - ---- - - - - ---- --- ---- - - ----------- - 24 Priapulida ------ ---- - - - - -- - - -- - ------ - -- ------ 24 Nematoda - -- - --- --- -- - -- --- - -- --- ---- -- - - -- -- 24 Mollusca ------------- --- --------------- ------ 24 Sipunculida ---------- --- ------------ ---- -- --- - 46 Echiurida ------ - --- - - - - - --- --- - -- --- - -- - - --- -

06 Pavia.Pmd
Bollettino della Società Paleontologica Italiana, 45 (2-3), 2006, 217-226. Modena, 15 gennaio 2007217 Lissoceras monachum (Gemmellaro), a ghost Ammonitida of the Tethyan Bathonian Giulio PAVIA G. Pavia, Dipartimento di Scienze della Terra, via Valperga Caluso 35, I-10124 Torino (Italy); [email protected] KEY-WORDS - Systematics, Ammonitida, Lissoceras, Bathonian, Tethys. ABSTRACT - L. monachum has been frequently recorded in the Upper Bajocian and Lower Bathonian, but references in literature differ in morphological details as they are based on a juvenile and poorly-preserved holotype. In addition, the type of L. monachum comes from a bed affected by taphonomical condensation mixing fossils from Early and Middle Bathonian times, i.e. the biochronological meaning of Gemmellaro’s taxon cannot be specified with biostratigraphic criteria from the type-locality. These references are here revised and refused on the basis of two topotypes recently sampled at Monte Erice in western Sicily, which allow a precise morphological definition of L. monachum by means of architectural and sutural characteristics. The only acceptable biostratigraphic datum comes from a Lower Bathonian specimen from southern France. The differences from L. ferrifex, L. magnum, and L. ventriplanum are discussed. Finally, the suture-line of topotype PU111502 of L. monachum and those specimens from the Upper Bajocian of the Venetian Alps, here named L. aff. monachum, support the phyletic relationships between L. ferrifex and L. magnum through transitional forms of L. monachum aged for the Tethyan Early Bathonian. RIASSUNTO - [Lissoceras monachum (Gemmellaro), un Ammonitida fantasma del Batoniano Tetideo] - L’ammonite Lissoceras monachum, della famiglia medio- e tardo-giurassica Lissoceratidae, risulta ripetutamente citata nella letteratura sistematica relativa al Baiociano superiore e al Batoniano inferiore. -

Download PDF ( Final Version , 5Mb )
Nautiloïden en ammonieten uit Maas- en Rijngrind in Limburg en zuidelijk Gelderland Philip+J. Hoedemaeker John+W.M. Jagt in Algemeen zandgroeve Leccius de Ridderop de Grebbeberg (Rhenen, Gelderland), en heeft als grootste diameter74 mm; de navel de meet 23 mm en windingshoogte en -breedte zijn 32 en ca. in menige verzameling te vinden zijn nautili- 34 mm. Burger refereert ook aan eerdere gemelde vondsten HOEWELden en ammonieten van paleozoïsche en mesozoïsche van fossielen van Oxfordien-ouderdom in grindpakketten in (Jura-Krijt) ouderdom uit Maas- en Rijngrindpakketten Midden-Nederland, en gaatervan uit dat deze, net als de door in Limburg en Gelderlander altijd bekaaid van afgekomen. Uit- hem gemelde ammoniet, duidelijke Maascomponenten zijn. De soort C. zonderingen op die regel zijn Teloceras soorten, waaraan een lillooetenselijkt echter van Noord-Amerikaanse aantalartikelen tussen 1970en de dag van vandaag is gewijd, (Canadese) origine te zijn en is, voor zover wij hebben kunnen Goniatitida26. Keienboek290 en In Het wordt gewag gemaakt nagaan, nooit uit noordwest Europa gemeld. Een vergelijk 122,226,230 van het voorkomen in Maasgrind van Teloceras als ‘de meest met recente literaturuur doet vermoeden dat het verbreide ammoniet in het Nederlandse zwerfsteenmateriaal; exemplaar uit Rhenen toch nauwer aansluit bij C. cordatum Willems kon uit vele kenmerkend (1960, 1970) Zuid-Limburg 20 exemplaren op- (J. Sowerby, 1813) en zijn verwanten, voor het sommen, maar ook elders langs de Maas (Mill, Mook), zowel vroeg-Oxfordien. als in de Achterhoek zijn fragmenten en gave exemplaren van deze fraaie in Materiaal hier Dogger-ammoniet verkiezelde toestand aange- voorgesteld is in hoofdzaak afkomstig uit de troffen.’ Als herkomst werd collecties het Nederlands genoteerd, ‘bij Mezières aan de van Centrum voor Biodiversiteit in komen dezelfde het Maas Frankrijk precies verkiezelde ammo- (Naturalis, Leiden) en Natuurhistorisch Museum Het nieten nog in hetvaste gesteentevoor.’ Als laatste noemt Van Maastricht. -

First Record and Correlation Value of Aulacostephanus Cf. Subundorae (PAVLOW) (Ammonoidea, Upper Jurassic) from SW Germany
N. Jb. Geol. Paläont. Mh. 2005 (2) 65-82 Stuttgart, Februar 2005 First record and correlation value of Aulacostephanus cf. subundorae (PAVLOW) (Ammonoidea, Upper Jurassic) from SW Germany Günter Schweigert and Lothar H. Vallon, Stuttgart With 7 figures and 1 table SCHWEIGERT, G. & VALLON, L. H. (2005): First record and correlation value of Aulacostephanus cf. subundorae (PAVLOW) (Ammonoidea, Upper Jurassic) from SW Germany. - N. Jb. Geol. Paläont. Mh., 2005: 65-82; Stuttgart. Abstract: Aulacostephanus cf. subundorae (PAVLOW) is recorded lor the first time from the Upper Jurassic Brenztaltrümmerkalk Member in Eastern Swabia (SW Germany). Together with Gravesia iritis (D'OKBIGNY), this ammonite species of Subboreal origin is indicative of the youngest Late Kimmeridgian Autissiodorensis Zone. The co-occurring ammonites from this formation have a Submediterranean origin. Hence, a correlation is possible between the Subboreal and the Submedi- terranean zonation around the Kimmeridgian/Tithonian boundary. Zusammenfassung: Die subboreale Ammonitenart Aulacostephanus cf. subundorae (PAVLOW) wird erstmals aus der Brenztaltrümmerkalk-Subformation im Oberjura der östlichen Schwäbischen Alb nachgewiesen. Zusammen mit Gravesia irius (D'ORBIGNY) kennzeichnet die Art die Autissiodorensis-Zone des jüngsten Kim- meridgium. Die begleitende Ammonitenfauna ist der Submediterranen Faunen- provinz zuzurechnen. Hierdurch wird eine Korrelation zwischen der subborealen und der submediterranen Zonengliederung im Kimmeridgium/Tithonium-Grenzbereich ermöglicht. 0028-3630/05/2005-0065 $ 4.50 © 2005 E. Schweizerbart'sche Verlagsbuchhandlung, D-70176 Stuttgart 66 G. Schweigert and L. H. Vallon 1. Introduction In the eastern part of Swabia, the Brenztaltrümmerkalk is a locally developed bioclastic deposit in the higher part of the Upper Jurassic. The Brenztal- trümmerkalk has a maximum thickness of about 100 meters and is mainly restricted to the near surroundings of Heidenheim and Schnaitheim, where it was quarried for several centuries as an important building stone. -

Kimmeridgian (Late Jurassic) Cold-Water Idoceratids (Ammonoidea) from Southern Coahuila, Northeastern Mexico, Associated with Boreal Bivalves and Belemnites
REVISTA MEXICANA DE CIENCIAS GEOLÓGICAS Kimmeridgian cold-water idoceratids associated with Boreal bivalvesv. 32, núm. and 1, 2015, belemnites p. 11-20 Kimmeridgian (Late Jurassic) cold-water idoceratids (Ammonoidea) from southern Coahuila, northeastern Mexico, associated with Boreal bivalves and belemnites Patrick Zell* and Wolfgang Stinnesbeck Institute for Earth Sciences, Heidelberg University, Im Neuenheimer Feld 234, 69120 Heidelberg, Germany. *[email protected] ABSTRACT et al., 2001; Chumakov et al., 2014) was followed by a cool period during the late Oxfordian-early Kimmeridgian (e.g., Jenkyns et al., Here we present two early Kimmeridgian faunal assemblages 2002; Weissert and Erba, 2004) and a long-term gradual warming composed of the ammonite Idoceras (Idoceras pinonense n. sp. and trend towards the Jurassic-Cretaceous boundary (e.g., Abbink et al., I. inflatum Burckhardt, 1906), Boreal belemnites Cylindroteuthis 2001; Lécuyer et al., 2003; Gröcke et al., 2003; Zakharov et al., 2014). cuspidata Sachs and Nalnjaeva, 1964 and Cylindroteuthis ex. gr. Palynological data suggest that the latest Jurassic was also marked by jacutica Sachs and Nalnjaeva, 1964, as well as the Boreal bivalve Buchia significant fluctuations in paleotemperature and climate (e.g., Abbink concentrica (J. de C. Sowerby, 1827). The assemblages were discovered et al., 2001). in inner- to outer shelf sediments of the lower La Casita Formation Upper Jurassic-Lower Cretaceous marine associations contain- at Puerto Piñones, southern Coahuila, and suggest that some taxa of ing both Tethyan and Boreal elements [e.g. ammonites, belemnites Idoceras inhabited cold-water environments. (Cylindroteuthis) and bivalves (Buchia)], were described from numer- ous localities of the Western Cordillera belt from Alaska to California Key words: La Casita Formation, Kimmeridgian, idoceratid ammonites, (e.g., Jeletzky, 1965), while Boreal (Buchia) and even southern high Boreal bivalves, Boreal belemnites.