Insulin Inhibits Leptin Receptor Signalling in HEK293 Cells at the Level of Janus Kinase-2: a Potential Mechanism for Hyperinsulinaemia-Associated Leptin Resistance
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

DNA Damage and the Activation of the P53 Pathway Mediate Alterations in Metabolic and Secretory Functions of Adipocytes
3062 Diabetes Volume 65, October 2016 Bastien Vergoni,1,2 Pierre-Jean Cornejo,1,2 Jérôme Gilleron,1,2 Mansour Djedaini,1,2 Franck Ceppo,1,2 Arnaud Jacquel,2,3 Gwennaelle Bouget,1,2 Clémence Ginet,1,2 Teresa Gonzalez,1,2,4 Julie Maillet,5,6,7 Véronique Dhennin,5,6,7 Marie Verbanck,5,6,7 Patrick Auberger,2,3 Philippe Froguel,5,6,7,8 Jean-François Tanti,1,2 and Mireille Cormont1,2 DNA Damage and the Activation of the p53 Pathway Mediate Alterations in Metabolic and Secretory Functions of Adipocytes Diabetes 2016;65:3062–3074 | DOI: 10.2337/db16-0014 Activation of the p53 pathway in adipose tissue contributes Insulin resistance is associated with obesity and is a major to insulin resistance associated with obesity. However, the risk factor for type 2 diabetes. Adipose tissue (AT) plays mechanisms of p53 activation and the effect on adipocyte an important role in glucose and lipid homeostasis (1,2), functions are still elusive. Here we found a higher level of and AT dysfunction associated with obesity has emerged DNA oxidation and a reduction in telomere length in adipose as a critical event for the development of insulin resis- tissueofmicefedahigh-fatdietandanincreaseinDNA tance. Low-grade inflammation and oxidative and hypoxic damage and activation of the p53 pathway in adipocytes. stresses both develop in obese AT and are involved in the Interestingly, hallmarks of chronic DNA damage are visible alteration of adipocyte functions (3–6). Therefore, activa- at the onset of obesity. Furthermore, injection of lean mice tion of stress-signaling pathways in adipocytes is critically with doxorubicin, a DNA damage-inducing drug, increased involved in AT dysfunction and insulin resistance (7–10). -

Selective Insulin Signaling Through a and B Insulin Receptors Regulates Transcription of Insulin and Glucokinase Genes in Pancreatic  Cells
Molecular Cell, Vol. 7, 559±570, March, 2001, Copyright 2001 by Cell Press Selective Insulin Signaling through A and B Insulin Receptors Regulates Transcription of Insulin and Glucokinase Genes in Pancreatic  Cells Barbara Leibiger,*§ Ingo B. Leibiger,*§k ceptors as the primary target, include signaling via mito- Tilo Moede,* Sabine Kemper,* gen-activated protein (MAP) kinases and phosphoinosi- Rohit N. Kulkarni,² C. Ronald Kahn,² tol-3 kinase (PI3K). The insulin receptor (IR), the first Lina Moitoso de Vargas,³ and Per-Olof Berggren* step in these cascades, exists in two isoforms as a result *The Rolf Luft Center for Diabetes Research of alternative mRNA splicing of the 11th exon of the insulin Department of Molecular Medicine proreceptor transcript (Seino et al., 1989). The A type Karolinska Institutet (IR-A), or Ex11Ϫ (Ullrich et al., 1985), lacks whereas the S-171 76 Stockholm B type (IR-B), or Ex11ϩ (Ebina et al., 1985), contains Sweden the respective sequence coding for 12 amino acids in ² Research Division the C terminus of the ␣ chain of the receptor. To date, Joslin Diabetes Center and no insulin-induced effect has been reported that dis- Department of Medicine criminates signaling via A- and B-type receptors. In fact, Harvard Medical School the functional significance of these IR isoforms remains Boston, Massachusetts 02215 unclear. ³ Department of Medicine Recent studies have shown that the insulin-producing New England Medical Center and pancreatic  cell is a target for insulin action, with insulin Tufts University School of Medicine effects on transcription, translation, Ca2ϩ flux, and exo- Boston, Massachusetts 02111 cytosis (Leibiger et al., 1998a, 2000; Xu and Rothenberg, 1998; Xu et al., 1998; Aspinwall et al., 1999; Kulkarni et al., 1999a). -

Insulin–Insr Signaling Drives Multipotent Progenitor Differentiation Toward Lymphoid Lineages
Article Insulin–InsR signaling drives multipotent progenitor differentiation toward lymphoid lineages Pengyan Xia,1* Shuo Wang,1* Ying Du,1 Guanling Huang,1,2 Takashi Satoh,3 Shizuo Akira,3 and Zusen Fan1 1Key Laboratory of Infection and Immunity of CAS, Institute of Biophysics, Chinese Academy of Sciences, Beijing 100101, China 2University of Chinese Academy of Sciences, Beijing 100049, China 3Department of Host Defense, Research Institute for Microbial Diseases (RIMD), Osaka University, Suita, Osaka 565-0871, Japan The lineage commitment of HSCs generates balanced myeloid and lymphoid populations in hematopoiesis. However, the un- derlying mechanisms that control this process remain largely unknown. Here, we show that insulin–insulin receptor (InsR) signaling is required for lineage commitment of multipotent progenitors (MPPs). Deletion of Insr in murine bone marrow causes skewed differentiation of MPPs to myeloid cells. mTOR acts as a downstream effector that modulates MPP differentiation. mTOR activates Stat3 by phosphorylation at serine 727 under insulin stimulation, which binds to the promoter of Ikaros, lead- ing to its transcription priming. Our findings reveal that the insulin–InsR signaling drives MPP differentiation into lymphoid lineages in early lymphopoiesis, which is essential for maintaining a balanced immune system for an individual organism. Hematopoiesis is the process of producing all compo- Insulin, as the primary anabolic hormone, modulates a nents of the blood system from hematopoietic stem cells variety of physiological processes, including growth, differ- (HSCs; Naik et al., 2013; Mendelson and Frenette, 2014; entiation, apoptosis, and synthesis and breakdown of lipid, Walter et al., 2015). HSCs are quiescent, self-renewable pro- protein, and glucose (Samuel and Shulman, 2012). -

PSM, a Mediator of PDGF-BB-, IGF-I-, and Insulin-Stimulated Mitogenesis
Oncogene (2000) 19, 39 ± 50 ã 2000 Macmillan Publishers Ltd All rights reserved 0950 ± 9232/00 $15.00 www.nature.com/onc PSM, a mediator of PDGF-BB-, IGF-I-, and insulin-stimulated mitogenesis Heimo Riedel*,1,2, Nasim Yousaf1, Yuyuan Zhao4, Heping Dai1,3, Youping Deng1 and Jian Wang1 1Department of Biological Sciences, Wayne State University, Detroit, Michigan, MI 48202, USA; 2Member, Barbara Ann Karmanos Cancer Institute, Detroit, Michigan, MI 48201, USA PSM/SH2-B has been described as a cellular partner of Introduction the FceRI receptor, insulin receptor (IR), insulin-like growth factor-I (IGF-I) receptor (IGF-IR), and nerve The platelet-derived growth factor (PDGF) family growth factor receptor (TrkA). A function has been represents key mitogenic and chemotactic factors that proposed in neuronal dierentiation and development but have been exploited by the Simian sarcoma virus v-sis its role in other signaling pathways is still unclear. To oncogene which results in altered PDGF expression further elucidate the physiologic role of PSM we have (Water®eld et al., 1983). Normal cellular PDGF identi®ed additional mitogenic receptor tyrosine kinases appears in three distinct isoforms as any combination as putative PSM partners including platelet-derived of the PDGF A and B chains which act in paracrine growth factor (PDGF) receptor (PDGFR) beta, hepato- and autocrine mechanisms (Heldin, 1993). PDGF cyte growth factor receptor (Met), and ®broblast growth receptor (PDGFR) signal transduction appears to be factor receptor. We have mapped Y740 as a site of a largely membrane delineated event (Hughes et al., PDGFR beta that is involved in the association with 1996). -

Investigation of the Underlying Hub Genes and Molexular Pathogensis in Gastric Cancer by Integrated Bioinformatic Analyses
bioRxiv preprint doi: https://doi.org/10.1101/2020.12.20.423656; this version posted December 22, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. Investigation of the underlying hub genes and molexular pathogensis in gastric cancer by integrated bioinformatic analyses Basavaraj Vastrad1, Chanabasayya Vastrad*2 1. Department of Biochemistry, Basaveshwar College of Pharmacy, Gadag, Karnataka 582103, India. 2. Biostatistics and Bioinformatics, Chanabasava Nilaya, Bharthinagar, Dharwad 580001, Karanataka, India. * Chanabasayya Vastrad [email protected] Ph: +919480073398 Chanabasava Nilaya, Bharthinagar, Dharwad 580001 , Karanataka, India bioRxiv preprint doi: https://doi.org/10.1101/2020.12.20.423656; this version posted December 22, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. Abstract The high mortality rate of gastric cancer (GC) is in part due to the absence of initial disclosure of its biomarkers. The recognition of important genes associated in GC is therefore recommended to advance clinical prognosis, diagnosis and and treatment outcomes. The current investigation used the microarray dataset GSE113255 RNA seq data from the Gene Expression Omnibus database to diagnose differentially expressed genes (DEGs). Pathway and gene ontology enrichment analyses were performed, and a proteinprotein interaction network, modules, target genes - miRNA regulatory network and target genes - TF regulatory network were constructed and analyzed. Finally, validation of hub genes was performed. The 1008 DEGs identified consisted of 505 up regulated genes and 503 down regulated genes. -

Structure of the Tie2 RTK Domain: Self-Inhibition by the Nucleotide Binding Loop, Activation Loop, and C-Terminal Tail
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Elsevier - Publisher Connector Structure, Vol. 8, 1105±1113, November, 2000, 2000 Elsevier Science Ltd. All rights reserved. PII S0969-2126(00)00516-5 Structure of the Tie2 RTK Domain: Self-Inhibition by the Nucleotide Binding Loop, Activation Loop, and C-Terminal Tail Lisa M. Shewchuk,* Anne M. Hassell, Byron Ellis, binding domain and an intracellular kinase domain. Binding of W. D. Holmes, Roderick Davis, Earnest L. Horne, extracellular ligand is believed to promote dimerization, which Sue H. Kadwell, David D. McKee, leads to autophosphorylation and activation of the kinase do- and John T. Moore main [reviewed in 15]. Stringent regulation of the phosphoryla- GlaxoWellcome tion state and activity of the Tie2 kinase domain is crucial to Research Triangle Park, North Carolina 27709 normal vasculature development and maintenance. Tie2 activ- ity is precisely regulated by the opposing actions of agonistic and antagonistic extracellular ligands [16±18]. Tie2 activation Summary requires autophosporylation in response to binding its agonists angiopoietin 1 (Ang1) and Ang4, whereas inactivation occurs Background: Angiogenesis, the formation of new vessels from in response to Ang2 and Ang3. Tie2 mutations, which result the existing vasculature, is a critical process during early devel- in ligand-independent and enhanced autophosphorylation, opment as well as in a number of disease processes. Tie2 (also cause hereditary venous malformations [19, 20]. Conversely, known as Tek) is an endothelium-specific receptor tyrosine transgenic mice that express a kinase-inactive form of Tie2 or kinase involved in both angiogenesis and vasculature mainte- Tie2 null mice die in utero due to defects in their microvascula- nance. -

High-Fat Diet-Induced Complement Activation Mediates Intestinal Inflammation and Neoplasia, Independent of Obesity Stephanie K
Published OnlineFirst August 17, 2016; DOI: 10.1158/1541-7786.MCR-16-0153 Metabolism Molecular Cancer Research High-Fat Diet-Induced Complement Activation Mediates Intestinal Inflammation and Neoplasia, Independent of Obesity Stephanie K. Doerner1, Edimara S. Reis2, Elaine S. Leung3, Justine S. Ko1, Jason D. Heaney4,5, Nathan A. Berger1,6, John D. Lambris2, and Joseph H. Nadeau1,3 Abstract Obesity and related metabolic disturbances are closely asso- and generation of C5a, which in turn induces the production of ciated with pathologies that represent a significant burden to proinflammatory cytokines and expression of proto-oncogenes. global health. Epidemiological and molecular evidence links Pharmacological and genetic targeting of the C5a receptor obesity and metabolic status with inflammation and increased reduced both inflammation and intestinal polyposis, suggest- risk of cancer. Here, using a mouse model of intestinal neo- ing the use of complement inhibitors for preventing diet- plasia and strains that are susceptible or resistant to diet- induced neoplasia. induced obesity, it is demonstrated that high-fat diet-induced inflammation, rather than obesity or metabolic status, is asso- Implications: This study characterizes the relations between diet ciated with increased intestinal neoplasia. The complement and metabolic conditions on risk for a common cancer and fragmentC5aactsasthetriggerforinflammation and intestinal identifies complement activation as a novel target for cancer tumorigenesis. High-fat diet induces complement activation prevention. Mol Cancer Res; 14(10); 953–65. Ó2016 AACR. Introduction Supporting a role for diet-induced obesity (DIO) in promoting inflammation and cancer, levels of IL6 and tumor burden are both Obesity is an increasingly important risk factor for many ob ob reduced following azoxymethane treatment of obese Lep / mice cancers (1, 2). -

Leptin As a Uremic Toxin Interferes with Neutrophil Chemotaxis
J Am Soc Nephrol 15: 2366–2372, 2004 Leptin as a Uremic Toxin Interferes with Neutrophil Chemotaxis LUCIANO OTTONELLO,* PAOLA GNERRE,* MARIA BERTOLOTTO,* MARINA MANCINI,* PATRIZIA DAPINO,* RODOLFO RUSSO,† GIACOMO GARIBOTTO,† TOMMASO BARRECA,* and FRANCO DALLEGRI* *Division of Internal Medicine and †Division of Nephrology, Department of Internal Medicine and Medical Specialties, University of Genoa Medical School, Genoa, Italy Abstract. Leptin is a pleiotropic molecule involved in energy Finally, the serum inhibitory activity can be effectively prevented homeostasis, hematopoiesis, inflammation, and immunity. Hypo- by immune depletion of leptin. The results also show, however, leptinemia characterizing starvation has been strictly related to that leptin by itself is endowed with chemotactic activity toward increased susceptibility to infection secondary to malnutrition. neutrophils. The two activities—inhibition of the cell response to Nevertheless, ESRD is characterized by high susceptibility to chemokines and stimulation of neutrophil migration—could be bacterial infection despite hyperleptinemia. Defects in neutrophils detected at similar concentrations. On the contrary, neutrophils 2ϩ play a crucial role in the infectious morbidity, and several uremic exposed to leptin did not display detectable [Ca ]i mobilization,  toxins that are capable of depressing neutrophil functions have oxidant production, or 2-integrin upregulation. The results dem- been identified. Only a few and contrasting reports about leptin onstrate that leptin is -
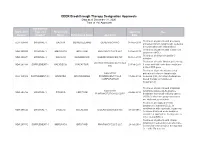
CDER Breakthrough Therapy Designation Approvals Data As of December 31, 2020 Total of 190 Approvals
CDER Breakthrough Therapy Designation Approvals Data as of December 31, 2020 Total of 190 Approvals Submission Application Type and Proprietary Approval Use Number Number Name Established Name Applicant Date Treatment of patients with previously BLA 125486 ORIGINAL-1 GAZYVA OBINUTUZUMAB GENENTECH INC 01-Nov-2013 untreated chronic lymphocytic leukemia in combination with chlorambucil Treatment of patients with mantle cell NDA 205552 ORIGINAL-1 IMBRUVICA IBRUTINIB PHARMACYCLICS LLC 13-Nov-2013 lymphoma (MCL) Treatment of chronic hepatitis C NDA 204671 ORIGINAL-1 SOVALDI SOFOSBUVIR GILEAD SCIENCES INC 06-Dec-2013 infection Treatment of cystic fibrosis patients age VERTEX PHARMACEUTICALS NDA 203188 SUPPLEMENT-4 KALYDECO IVACAFTOR 21-Feb-2014 6 years and older who have mutations INC in the CFTR gene Treatment of previously untreated NOVARTIS patients with chronic lymphocytic BLA 125326 SUPPLEMENT-60 ARZERRA OFATUMUMAB PHARMACEUTICALS 17-Apr-2014 leukemia (CLL) for whom fludarabine- CORPORATION based therapy is considered inappropriate Treatment of patients with anaplastic NOVARTIS lymphoma kinase (ALK)-positive NDA 205755 ORIGINAL-1 ZYKADIA CERITINIB 29-Apr-2014 PHARMACEUTICALS CORP metastatic non-small cell lung cancer (NSCLC) who have progressed on or are intolerant to crizotinib Treatment of relapsed chronic lymphocytic leukemia (CLL), in combination with rituximab, in patients NDA 206545 ORIGINAL-1 ZYDELIG IDELALISIB GILEAD SCIENCES INC 23-Jul-2014 for whom rituximab alone would be considered appropriate therapy due to other co-morbidities -

LCZ696) Compared to Valsartan Attenuates Hepatotoxicity in STZ-Induced Hyperglycemic Rats Faleh Alqahtani#, Mohamed Mohany#, Abdullah F Alasmari, Ahmed Z
Int. J. Med. Sci. 2020, Vol. 17 3098 Ivyspring International Publisher International Journal of Medical Sciences 2020; 17(18): 3098-3106. doi: 10.7150/ijms.49373 Research Paper Angiotensin II receptor Neprilysin inhibitor (LCZ696) compared to Valsartan attenuates Hepatotoxicity in STZ-induced hyperglycemic rats Faleh Alqahtani#, Mohamed Mohany#, Abdullah F Alasmari, Ahmed Z. Alanazi, Osamah M. Belali, Mohammed M Ahmed, and Salim S Al-Rejaie Department of Pharmacology and Toxicology, College of Pharmacy, King Saud University, P.O. Box 55760, Riyadh – 1145, Saudi Arabia. #These authors contributed equally to this work. Corresponding author: E-mail: [email protected]; Tel.: +966114677178. © The author(s). This is an open access article distributed under the terms of the Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0/). See http://ivyspring.com/terms for full terms and conditions. Received: 2020.06.11; Accepted: 2020.10.09; Published: 2020.10.22 Abstract Background and objectives: Although diabetic-induced hepatotoxicity is less common, it can be included in the list of target organ pathologies associated with diabetes. This study aimed to investigate the potential therapeutic role of sacubitril/valsartan (LCZ696) in modulating oxidative and inflammatory injuries and liver fibrosis in STZ-induced hyperglycemic rats in comparison to valsartan alone. Materials and Methods: Following the induction of diabetes using a single dose of streptozotocin (STZ), STZ-induced hyperglycemic animals were administered LCZ696 or valsartan for 6 weeks. Glucose, transaminases, lipid profile, tumor necrosis factor-alpha (TNF-α), interleukin 1 beta (IL-1β), and interleukin - 6 (IL-6), were estimated using the obtained serum. Oxidative stress biomarkers including thiobarbituric acid reactive substances (TBARS), glutathione (GSH), superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GPx), and glutathione S-transferase (GST) were measured in the liver homogenate. -

The Insulin and IGF-1 Receptors
The insulin and IGF-1 receptors Author: Pierre De Meyts Author title: MD, PHD, F.A.C.E. A bit of history The birth of the receptor concept dates back to the early work of John Newport Langley (1852-1925), a Cambridge physiologist, who postulated in 1905 that a "receptive substance" on the surface of skeletal muscle mediated the action of nicotine (1). At about the same time, Paul Ehrlich (1854-1915), a German immunologist who was the founding father of chemotherapy, came up with a "side chain theory" of cell receptors to explain the selectivity of immune reactions, winning him the Nobel Prize in Physiology or Medicine in 1908. His famous adage "Corpora non agunt nisi fixata" ("Substances do not act unless they are bound") is an elegant and concise early statement of the receptor theory (2). The receptor concept was put on more solid ground with the seminal 1948 paper of Raymond P. Ahlquist (1914-1983), an American pharmacologist of Swedish descent at the Medical College of Georgia, who proposed that the excitatory and inhibitory effects of adrenotropic agents were mediated by two separate receptors which he termed a and b (3). The receptors would however remain hypothetical entities until the late 60's, when direct methods to study their biochemistry were developed. The concept that insulin exerts its effects by acting at the membrane of target cells was proposed over 60 years ago by Rachmiel Levine (1910-1991), considered by many as one of the founding fathers of modern diabetology, then working at Walter Reese Hospital in Chicago. -

Five Mutant Alleles of the Insulin Receptor Gene in Patients with Genetic Forms of Insulin Resistance
Five mutant alleles of the insulin receptor gene in patients with genetic forms of insulin resistance. T Kadowaki, … , P Gorden, S I Taylor J Clin Invest. 1990;86(1):254-264. https://doi.org/10.1172/JCI114693. Research Article The nucleotide sequence was determined for all 22 exons of the insulin receptor gene from three patients with genetic syndromes associated with extreme insulin resistance. In all three patients, insulin resistance was caused by decreased insulin binding to the cell surface. The patient with leprechaunism (leprechaun/Winnipeg) came from a consanguineous pedigree and was homozygous for a missense mutation substituting arginine for His209 in the alpha-subunit of the insulin receptor. The other two patients were both compound heterozygotes with a nonsense mutation in one allele of the insulin receptor gene, and a missense mutation in the other allele. In the patient with the Rabson-Mendenhall syndrome (patient RM-1), the missense mutation substituted lysine for Asn15 in the alpha-subunit. In the patient with type A extreme insulin resistance (patient A-1), the missense mutation substituted serine for Asn462 in the alpha-subunit. Both nonsense mutations markedly reduced the levels of insulin receptor mRNA transcribed from the alleles with the nonsense mutation as compared to the transcripts from the other allele. The reduction in the level of mRNA would be predicted to greatly reduce the rate at which the truncated receptors would be synthesized. Furthermore, the truncated receptors would be severely impaired in their ability to mediate insulin action. Find the latest version: https://jci.me/114693/pdf Five Mutant Alleles of the Insulin Receptor Gene in Patients with Genetic Forms of Insulin Resistance Takashi Kadowaki,* Hiroko Kadowaki,* Matthew M.