High-Fat Diet-Induced Complement Activation Mediates Intestinal Inflammation and Neoplasia, Independent of Obesity Stephanie K
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Investigation of the Underlying Hub Genes and Molexular Pathogensis in Gastric Cancer by Integrated Bioinformatic Analyses
bioRxiv preprint doi: https://doi.org/10.1101/2020.12.20.423656; this version posted December 22, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. Investigation of the underlying hub genes and molexular pathogensis in gastric cancer by integrated bioinformatic analyses Basavaraj Vastrad1, Chanabasayya Vastrad*2 1. Department of Biochemistry, Basaveshwar College of Pharmacy, Gadag, Karnataka 582103, India. 2. Biostatistics and Bioinformatics, Chanabasava Nilaya, Bharthinagar, Dharwad 580001, Karanataka, India. * Chanabasayya Vastrad [email protected] Ph: +919480073398 Chanabasava Nilaya, Bharthinagar, Dharwad 580001 , Karanataka, India bioRxiv preprint doi: https://doi.org/10.1101/2020.12.20.423656; this version posted December 22, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. Abstract The high mortality rate of gastric cancer (GC) is in part due to the absence of initial disclosure of its biomarkers. The recognition of important genes associated in GC is therefore recommended to advance clinical prognosis, diagnosis and and treatment outcomes. The current investigation used the microarray dataset GSE113255 RNA seq data from the Gene Expression Omnibus database to diagnose differentially expressed genes (DEGs). Pathway and gene ontology enrichment analyses were performed, and a proteinprotein interaction network, modules, target genes - miRNA regulatory network and target genes - TF regulatory network were constructed and analyzed. Finally, validation of hub genes was performed. The 1008 DEGs identified consisted of 505 up regulated genes and 503 down regulated genes. -

Trends in Beverage Consumption and Related Demographic Factors and Obesity Among Korean Children and Adolescents
nutrients Article Trends in Beverage Consumption and Related Demographic Factors and Obesity among Korean Children and Adolescents Su Bin Hwang, SoHyun Park , Guang-Ri Jin, Jae Hyun Jung, Hyeon Ju Park, Su Hyun Lee, Sangah Shin * and Bog-Hieu Lee * Department of Food and Nutrition, Chung-Ang University, Gyeonggi-do 17546, Korea; [email protected] (S.B.H.); [email protected] (S.P.); [email protected] (G.-R.J.); [email protected] (J.H.J.); [email protected] (H.J.P.); [email protected] (S.H.L.) * Correspondence: [email protected] (S.S.); [email protected] (B.-H.L.); Tel.: +82-31-670-3259 (S.S.); +82-31-670-3276 (B.-H.L.) Received: 24 July 2020; Accepted: 28 August 2020; Published: 31 August 2020 Abstract: It is well known that reducing consumption of sugar is a global public health priority. Beverages were the primary source of total sugar intake from processed foods. However, there are few studies investigating the trend of beverage consumption among children and adolescents in Korea. We examined the overall trend in beverage consumption among 11,996 participants aged 10–18 years who were enrolled in the Korea National Health and Nutrition Examination Survey (KNHANES) (1998–2018). Further, we examined the effect of beverage types on beverage consumption-related demographic factors and obesity among 6121 participants using the recent 24 h dietary recall data (2010–2018) that captured the consumption of fruit and vegetable juices, soft drinks, milk and milk-based products and alcoholic beverages. Demographic characteristics, including sex, age, body mass index, household income level and residential area, were considered. -

Leptin As a Uremic Toxin Interferes with Neutrophil Chemotaxis
J Am Soc Nephrol 15: 2366–2372, 2004 Leptin as a Uremic Toxin Interferes with Neutrophil Chemotaxis LUCIANO OTTONELLO,* PAOLA GNERRE,* MARIA BERTOLOTTO,* MARINA MANCINI,* PATRIZIA DAPINO,* RODOLFO RUSSO,† GIACOMO GARIBOTTO,† TOMMASO BARRECA,* and FRANCO DALLEGRI* *Division of Internal Medicine and †Division of Nephrology, Department of Internal Medicine and Medical Specialties, University of Genoa Medical School, Genoa, Italy Abstract. Leptin is a pleiotropic molecule involved in energy Finally, the serum inhibitory activity can be effectively prevented homeostasis, hematopoiesis, inflammation, and immunity. Hypo- by immune depletion of leptin. The results also show, however, leptinemia characterizing starvation has been strictly related to that leptin by itself is endowed with chemotactic activity toward increased susceptibility to infection secondary to malnutrition. neutrophils. The two activities—inhibition of the cell response to Nevertheless, ESRD is characterized by high susceptibility to chemokines and stimulation of neutrophil migration—could be bacterial infection despite hyperleptinemia. Defects in neutrophils detected at similar concentrations. On the contrary, neutrophils 2ϩ play a crucial role in the infectious morbidity, and several uremic exposed to leptin did not display detectable [Ca ]i mobilization,  toxins that are capable of depressing neutrophil functions have oxidant production, or 2-integrin upregulation. The results dem- been identified. Only a few and contrasting reports about leptin onstrate that leptin is -
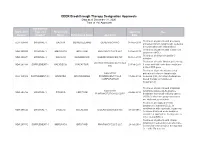
CDER Breakthrough Therapy Designation Approvals Data As of December 31, 2020 Total of 190 Approvals
CDER Breakthrough Therapy Designation Approvals Data as of December 31, 2020 Total of 190 Approvals Submission Application Type and Proprietary Approval Use Number Number Name Established Name Applicant Date Treatment of patients with previously BLA 125486 ORIGINAL-1 GAZYVA OBINUTUZUMAB GENENTECH INC 01-Nov-2013 untreated chronic lymphocytic leukemia in combination with chlorambucil Treatment of patients with mantle cell NDA 205552 ORIGINAL-1 IMBRUVICA IBRUTINIB PHARMACYCLICS LLC 13-Nov-2013 lymphoma (MCL) Treatment of chronic hepatitis C NDA 204671 ORIGINAL-1 SOVALDI SOFOSBUVIR GILEAD SCIENCES INC 06-Dec-2013 infection Treatment of cystic fibrosis patients age VERTEX PHARMACEUTICALS NDA 203188 SUPPLEMENT-4 KALYDECO IVACAFTOR 21-Feb-2014 6 years and older who have mutations INC in the CFTR gene Treatment of previously untreated NOVARTIS patients with chronic lymphocytic BLA 125326 SUPPLEMENT-60 ARZERRA OFATUMUMAB PHARMACEUTICALS 17-Apr-2014 leukemia (CLL) for whom fludarabine- CORPORATION based therapy is considered inappropriate Treatment of patients with anaplastic NOVARTIS lymphoma kinase (ALK)-positive NDA 205755 ORIGINAL-1 ZYKADIA CERITINIB 29-Apr-2014 PHARMACEUTICALS CORP metastatic non-small cell lung cancer (NSCLC) who have progressed on or are intolerant to crizotinib Treatment of relapsed chronic lymphocytic leukemia (CLL), in combination with rituximab, in patients NDA 206545 ORIGINAL-1 ZYDELIG IDELALISIB GILEAD SCIENCES INC 23-Jul-2014 for whom rituximab alone would be considered appropriate therapy due to other co-morbidities -

Human Induced Pluripotent Stem Cell–Derived Podocytes Mature Into Vascularized Glomeruli Upon Experimental Transplantation
BASIC RESEARCH www.jasn.org Human Induced Pluripotent Stem Cell–Derived Podocytes Mature into Vascularized Glomeruli upon Experimental Transplantation † Sazia Sharmin,* Atsuhiro Taguchi,* Yusuke Kaku,* Yasuhiro Yoshimura,* Tomoko Ohmori,* ‡ † ‡ Tetsushi Sakuma, Masashi Mukoyama, Takashi Yamamoto, Hidetake Kurihara,§ and | Ryuichi Nishinakamura* *Department of Kidney Development, Institute of Molecular Embryology and Genetics, and †Department of Nephrology, Faculty of Life Sciences, Kumamoto University, Kumamoto, Japan; ‡Department of Mathematical and Life Sciences, Graduate School of Science, Hiroshima University, Hiroshima, Japan; §Division of Anatomy, Juntendo University School of Medicine, Tokyo, Japan; and |Japan Science and Technology Agency, CREST, Kumamoto, Japan ABSTRACT Glomerular podocytes express proteins, such as nephrin, that constitute the slit diaphragm, thereby contributing to the filtration process in the kidney. Glomerular development has been analyzed mainly in mice, whereas analysis of human kidney development has been minimal because of limited access to embryonic kidneys. We previously reported the induction of three-dimensional primordial glomeruli from human induced pluripotent stem (iPS) cells. Here, using transcription activator–like effector nuclease-mediated homologous recombination, we generated human iPS cell lines that express green fluorescent protein (GFP) in the NPHS1 locus, which encodes nephrin, and we show that GFP expression facilitated accurate visualization of nephrin-positive podocyte formation in -

Is There a Link Between Different Types of Alcoholic Drinks And
International Journal of Environmental Research and Public Health Article Is There a Link between Different Types of Alcoholic Drinks and Obesity? An Analysis of 280,183 UK Biobank Participants Elif Inan-Eroglu 1,2, Lauren Powell 2, Mark Hamer 3, Gary O’Donovan 4 , Mitch J. Duncan 5,6 and Emmanuel Stamatakis 2,7,* 1 The Boden Collaboration for Obesity, Nutrition, Exercise & Eating Disorders, Faculty of Medicine and Health, The University of Sydney, Sydney NSW 2050, Australia; [email protected] 2 Prevention Research Collaboration, School of Public Health, Faculty of Medicine and Health, The University of Sydney, Sydney NSW 2050, Australia; [email protected] 3 Faculty of Medical Sciences, University College London, London WC1E 6BT, UK; [email protected] 4 Facultad de Medicina, Universidad de los Andes, Bogotá 57, Colombia; [email protected] 5 School of Medicine & Public Health; Faculty of Health and Medicine, The University of Newcastle, University Drive, Callaghan NSW 2308, Australia; [email protected] 6 Priority Research Centre for Physical Activity and Nutrition, The University of Newcastle, University Drive, Callaghan NSW 2308, Australia 7 Charles Perkins Centre Epidemiology Unit, The University of Sydney, Sydney NSW 2050, Australia * Correspondence: [email protected]; Tel.: +61-2-86271867 Received: 29 May 2020; Accepted: 13 July 2020; Published: 17 July 2020 Abstract: Understanding the associations between types of alcoholic drinks and adiposity has public health relevance, considering that adult overweight and obesity prevalence are increasing worldwide. We aimed to evaluate the association between overall alcohol consumption and types of alcohol drinks with markers of adiposity from the UK Biobank baseline data (n = 280,183, 48.3% female). -

Effect of Calorie Restriction and Exercise on Type 2
ПРИЛОЗИ. Одд. за мед. науки, XLII 1, 2021 МАНУ CONTRIBUTIONS. Sec. of Med. Sci., XLII 1, 2021 MASA 10.2478/prilozi-2021-0010 ISSN 1857-9345 UDC: 616.379-008.64:612.395.6 EFFECT OF CALORIE RESTRICTION AND EXERCISE ON TYPE 2 DIABETES Hira Shakoor1, Vasso Apostolopoulos2, Jack Feehan2, 3, Habiba Isse Ali1, Leila Cheikh Ismail 4, 5, Ayesha Salem Obaid S. Al Dhaheri1, Lily Stojanovska1, 2 1 Department of Nutrition and Health, College of Medicine and Health Sciences, United Arab Emirates, Al Ain, United Arab Emirates 2 Institute for Health and Sport,Victoria University, Melbourne, Australia 3 Department of Medicine-Western Health, Melbourne Medical School, The University of Melbourne, St. Albans, Australia 4 Department of Clinical Nutrition and Dietetics, College of Health Sciences, University of Sharjah, Sharjah, UAE 5 Nuffield Department of Women’s & Reproductive Health, University of Oxford, Oxford, UK Corresponding author: Lily Stojanovska, Department of Nutrition and Health, College of Medicine and Health Sciences, United Arab Emirates University, PO Box 15551, Al Ain, United Arab Emirates. Email:lily.stojanovaska@ uaeu.ac.ae Phone: +971525308064 ABSTRACT Type-2 diabetes (T2D) is a chronic condition, generally regarded as an irreversible, that is among the top 10 causes of death globally. The hallmark of T2D is hyperglycemia, which results from disturbances in insulin sensitivity, insulin secretion, β-cell dysfunction and insulin resistance. Several clinical and lifestyle factors are involved in the progression of T2D, such as obesity and physical inactivity. A high-calorie diet is the main contributor to the development of obesity, which results in T2D, as obesity or increased intra-abdominal adipose tissue is related to insulin resistance. -

Diet and Cardiometabolic Disease Dietary Trends and the Impact of Diet on Diabetes and Cardiovascular Disease in Northern Sweden
Diet and cardiometabolic disease Dietary trends and the impact of diet on diabetes and cardiovascular disease in northern Sweden Front cover: Sucktomten vallar lörpvålmar – illustration from a modern fairy tale describing the life of a benevolent goblin-sheppard in the woods of northern Sweden. This and all other illustrations are from the as yet unpublished book “Sucktomtens berättelser” by Tore ‘Rasp’ Hylander. Printed by permission of the author. • Copyright © 2007 by Benno Krachler ISBN 978-91-7264-354-3 Printed in Sweden by Print&Media Umeå university:2003462 Umeå 2007 2 To parents, teachers, tutors, colleagues, patients and all other friends with gratitude. 3 CONTENTS DIET AND CARDIOMETABOLIC DISEASE............................................. 1 DIETARY TRENDS AND THE IMPACT OF DIET ON DIABETES AND CARDIOVASCULAR DISEASE IN NORTHERN SWEDEN........... 1 ABSTRACT-ENGLISH............................................................................................................................................ 6 SAMMANFATTNING PÅ SVENSKA (ABSTRACT-SWEDISH)....................................................................................... 8 LIST OF PAPERS.................................................................................................................................................. 10 ABBREVIATIONS ............................................................................................................................................... 11 1 INTRODUCTION.................................................................................... -

Obesity and Sex Affect the Immune Responses to Tick-Borne Encephalitis Booster Vaccination
ORIGINAL RESEARCH published: 27 May 2020 doi: 10.3389/fimmu.2020.00860 Obesity and Sex Affect the Immune Responses to Tick-Borne Encephalitis Booster Vaccination Erika Garner-Spitzer 1*†, Eva-Maria Poellabauer 1†, Angelika Wagner 1, Angela Guzek 1, Ines Zwazl 1, Claudia Seidl-Friedrich 1, Christoph J. Binder 2, Karin Stiasny 3, Michael Kundi 4 and Ursula Wiedermann 1* 1 Institute of Specific Prophylaxis and Tropical Medicine, Medical University of Vienna, Vienna, Austria, 2 Department for Laboratory Medicine, Medical University Vienna, Vienna, Austria, 3 Center of Virology, Medical University Vienna, Vienna, Edited by: Austria, 4 Center for Public Health, Medical University Vienna, Vienna, Austria Karl Ljungberg, Eurocine Vaccines AB, Sweden Reviewed by: Obesity has dramatically increased over the last 30 years and reaches according to World Liubov I. Kozlovskaya, Health Organization dimensions of a global epidemic. The obesity-associated chronic Chumakov Federal Scientific Center for Research and Development of low-level inflammation contributes to severe comorbidities and directly affects many Immune-and- Biological Products immune cells leading to immune dysfunction and increased susceptibility to infections. (RAS), Russia Thus, prophylaxis against vaccine-preventable diseases is crucial, yet the responsiveness Galina Grigirievna Karganova, Chumakov Institute of Poliomyelitis to several vaccines is unclear under obesity. In order to assess the responsiveness and Viral Encephalitides (RAS), Russia to tick-borne encephalitis (TBE) vaccine, we revaccinated 37 obese individuals and *Correspondence: 36 normal-weight controls with a licensed TBE vaccine. Metabolic, hormonal, and Erika Garner-Spitzer [email protected] immunologic profiles along with vaccine-specific humoral and cellular immune responses Ursula Wiedermann were evaluated in sera and peripheral blood mononuclear cells (PBMCs) before, 1 week, [email protected] 4 weeks, and 6 months after TBE booster. -

Insulin and Leptin As Adiposity Signals
Insulin and Leptin as Adiposity Signals STEPHEN C. BENOIT,DEBORAH J. CLEGG,RANDY J. SEELEY, AND STEPHEN C. WOODS Department of Psychiatry, University of Cincinnati Medical Center, Cincinnati, Ohio 45267 ABSTRACT There is now considerable consensus that the adipocyte hormone leptin and the pancreatic hormone insulin are important regulators of food intake and energy balance. Leptin and insulin fulfill many of the requirements to be putative adiposity signals to the brain. Plasma leptin and insulin levels are positively correlated with body weight and with adipose mass in particular. Furthermore, both leptin and insulin enter the brain from the plasma. The brain expresses both insulin and leptin receptors in areas important in the control of food intake and energy balance. Consistent with their roles as adiposity signals, exogenous leptin and insulin both reduce food intake when administered locally into the brain in a number of species under different experimental paradigms. Additionally, central administration of insulin antibodies increases food intake and body weight. Recent studies have demonstrated that both insulin and leptin have additive effects when administered simulta- neously. Finally, we recently have demonstrated that leptin and insulin share downstream neuropep- tide signaling pathways. Hence, insulin and leptin provide important negative feedback signals to the central nervous system, proportional to peripheral energy stores and coupled with catabolic circuits. I. Overview When maintained on an ad libitum diet, most animals — including humans — are able to precisely match caloric intake with caloric expenditure, resulting in relatively stable energy stores as adipose tissue (Kennedy, 1953; Keesey, 1986). Growing emphasis has been placed on the role of the central nervous system (CNS) in controlling this precision of energy homeostasis. -

High-Fat Diet and Age-Dependent Effects of Iga-Bearing Cell Populations in the Small Intestinal Lamina Propria in Mice
International Journal of Molecular Sciences Communication High-Fat Diet and Age-Dependent Effects of IgA-Bearing Cell Populations in the Small Intestinal Lamina Propria in Mice Yuta Sakamoto 1,2 , Masatoshi Niwa 3, Ken Muramatsu 4 and Satoshi Shimo 5,* 1 Department of Physical Therapy, Faculty of Health Sciences, Health Science University, 7187 Kodachi, Fujikawaguchiko-machi, Minamitsuru-gun, Yamanashi 401-0380, Japan; [email protected] 2 Graduate School of Health Sciences, Kyorin University, 5-4-1 Shimorenjaku, Mitaka-shi, Tokyo 181-8612, Japan 3 Department of Occupational Therapy, Faculty of Health Sciences, Kyorin University, 5-4-1 Shimorenjaku, Mitaka-shi, Tokyo 181-8612, Japan; [email protected] 4 Department of Physical Therapy, Faculty of Health Sciences, Kyorin University, 5-4-1 Shimorenjaku, Mitaka-shi, Tokyo 181-8612, Japan; [email protected] 5 Department of Occupational Therapy, Faculty of Health Sciences, Health Science University, 7187 Kodachi, Fujikawaguchiko-machi, Minamitsuru-gun, Yamanashi 401-0380, Japan * Correspondence: [email protected]; Tel.: +81-555-83-5200 Abstract: Several studies highlighted that obesity and diabetes reduce immune function. However, changes in the distribution of immunoglobins (Igs), including immunoglobulin-A (IgA), that have an important function in mucosal immunity in the intestinal tract, are unclear. This study aimed to investigate the impaired immune functions in the context of a diet-induced obese murine model via the assessment of the Igs in the intestinal villi. We used mice fed a high-fat diet (HFD) from four to 12 or 20 weeks of age. The distributions of IgA, IgM, and IgG1 were observed by immunohistochemistry. -

Relevance of Serum Leptin and Leptin-Receptor Concentrations in Critically Ill Patients
Hindawi Publishing Corporation Mediators of Inflammation Volume 2010, Article ID 473540, 9 pages doi:10.1155/2010/473540 Clinical Study Relevance of Serum Leptin and Leptin-Receptor Concentrations in Critically Ill Patients Alexander Koch,1 Ralf Weiskirchen,2 Henning W. Zimmermann,1 Edouard Sanson,1 Christian Trautwein,1 and Frank Tacke1 1 Department of Medicine III, RWTH-University Hospital Aachen, Pauwelsstraße 30, 52074 Aachen, Germany 2 Institute of Clinical Chemistry and Pathobiochemistry, RWTH-University Hospital Aachen, Pauwelsstraße 30, 52074 Aachen, Germany Correspondence should be addressed to Alexander Koch, [email protected] Received 10 March 2010; Accepted 4 May 2010 Academic Editor: Oreste Gualillo Copyright © 2010 Alexander Koch et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. The adipocyte-derived cytokine leptin was implicated to link inflammation and metabolic alterations. We investigated the potential role of leptin components in critically ill patients, because systemic inflammation, insulin resistance, and hyperglycemia are common features of critical illness. Upon admission to Medical Intensive Care Unit (ICU), free leptin and soluble leptin-receptor serum concentrations were determined in 137 critically ill patients (95 with sepsis, 42 without sepsis) and 26 healthy controls. Serum leptin or leptin-receptor did not differ between patients or controls and were independent of sepsis. However, serum leptin was closely associated with obesity and diabetes and clearly correlated with markers of metabolism and liver function. Leptin- receptor was an unfavourable prognostic indicator, associated with mortality during three years follow-up.