Pollen Diagrams from Wolin Island (Nw Poland)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Pomerania in the Medieval and Renaissance Cartography – from the Cottoniana to Eilhard Lubinus
Pomerania in the Medieval and Renaissance Cartography… STUDIA MARITIMA, vol. XXXIII (2020) | ISSN 0137-3587 | DOI: 10.18276/sm.2020.33-04 Adam Krawiec Faculty of Historical Studies Adam Mickiewicz University, Poznań [email protected] ORCID: 0000-0002-3936-5037 Pomerania in the Medieval and Renaissance Cartography – from the Cottoniana to Eilhard Lubinus Keywords: Pomerania, Duchy of Pomerania, medieval cartography, early modern cartography, maritime cartography The following paper deals with the question of the cartographical image of Pomer- ania. What I mean here are maps in the modern sense of the word, i.e. Graphic rep- resentations that facilitate a spatial understanding of things, concepts, conditions, processes, or events in the human world1. It is an important reservation because the line between graphic and non-graphic representations of the Earth’s surface in the Middle Ages was sometimes blurred, therefore the term mappamundi could mean either a cartographic image or a textual geographical description, and in some cases it functioned as an equivalent of the modern term “Geography”2. Consequently, there’s a tendency in the modern historiography to analyze both forms of the geographical descriptions together. However, the late medieval and early modern developments in the perception and re-constructing of the space led to distinguishing cartography as an autonomous, full-fledged discipline of knowledge, and to the general acceptance of the map in the modern sense as a basic form of presentation of the world’s surface. Most maps which will be examined in the paper were produced in this later period, so it seems justified to analyze only the “real” maps, although in a broader context of the geographical imaginations. -

University of Auckland Research Repository, Researchspace
Libraries and Learning Services University of Auckland Research Repository, ResearchSpace Copyright Statement The digital copy of this thesis is protected by the Copyright Act 1994 (New Zealand). This thesis may be consulted by you, provided you comply with the provisions of the Act and the following conditions of use: • Any use you make of these documents or images must be for research or private study purposes only, and you may not make them available to any other person. • Authors control the copyright of their thesis. You will recognize the author's right to be identified as the author of this thesis, and due acknowledgement will be made to the author where appropriate. • You will obtain the author's permission before publishing any material from their thesis. General copyright and disclaimer In addition to the above conditions, authors give their consent for the digital copy of their work to be used subject to the conditions specified on the Library Thesis Consent Form and Deposit Licence. Sauerkraut and Salt Water: The German-Tongan Diaspora Since 1932 Kasia Renae Cook A thesis submitted in fulfilment of the requirements for the degree of Doctor of Philosophy in German, the University of Auckland, 2017. Abstract This is a study of individuals of German-Tongan descent living around the world. Taking as its starting point the period where Germans in Tonga (2014) left off, it examines the family histories, self-conceptions of identity, and connectedness to Germany of twenty-seven individuals living in New Zealand, the United States, Europe, and Tonga, who all have German- Tongan ancestry. -

Spatial Analysis of Plant Species Distribution in Midfield Ponds in an Agriculturally Intense Area
Pol. J. Environ. Stud. Vol. 21, No. 4 (2012), 871-877 Original Research Spatial Analysis of Plant Species Distribution in Midfield Ponds in an Agriculturally Intense Area Renata Gamrat1*, Małgorzata Gałczyńska2, Krzysztof Pacewicz3 1Department of Protection and Environmental Management, 2Department of General and Ecological Chemistry, 3Department of Agronomy, West Pomeranian University of Technology, Słowackiego 17, 71-434 Szczecin, Poland Received: 3 November 2011 Accepted: 22 March 2011 Abstract In an agricultural landscape small midfield ponds fulfill biocenotic and physiocenotic functions. A variety of species settle in areas of midfield ponds. The aim of our studies was to determine whether the occurrence of aquatic and marsh species in the studied bodies of water is a random effect or a result of spatial autocorrelation. On the basis of conclusions from ESDA, the hypothesis of spatial randomness can be reject- ed, which opens the way for searching spatial regimes. The review analysis of the spatial data (ESDA) with the use of join-count statistics showed that there is a positive spatial correlation for the midfield ponds in the agricultural area of the Pyrzyce-Stargard Plain in the distribution of ten aquatic and rush species for a distance of 1,250 m. The results of statistical analysis (ESDA) can constitute the basis for the choice and protection of midfield ponds as stop islands fulfilling functions of “mini” ecological corridors in spreading plant species. Keywords: ponds, plant species, distribution, spatial autocorrelation, lattice model Introduction including anthropogenic changes of flora, and the midfield ponds of small surface and depth undergo the processes of Western Pomerania includes numerous midfield ponds. -

Korekta Harmonogramu. Kwiecień 2021 R
HARMONOGRAM PRZETARGÓW w OT KOWR SZCZECIN na m-c KWIECIEŃ-2021 r. - KOREKTA www.kowr.gov.pl/nieruchomosci/harmonogramy telefon informacyjny: 91 81-44-223 KOWR OT w Szczecinie, ul. Bronowicka 41, 71-012 Szczecin, tel. 91 81 44 200, email: [email protected] Sekcja Zamiejscowa w Stargardzie, ul. Czarnieckiego 16, 73-110 Stargard, tel. 91 578 85 50, email: [email protected] Sekcja Zamiejscowa w Reczu, ul. Srebrna 22, 73-210 Recz, tel. 95 769 12 10, email: [email protected] Sekcja Zamiejscowa w Nowogardzie, ul. Główna 3, 72 – 200 Nowogard, tel. 91 578 93 70, email: [email protected] Sekcja Zamiejscowa w Gryficach, ul. Piłsudskiego 18, 72-300 Gryfice, tel. 91 384 77 10, email: [email protected] Sekcja Zamiejscowa w Pyrzycach, ul. Warszawska 95, 74-200 Pyrzyce, tel. 91 817 23 46, email: [email protected] POWIERZCHNIA POŁOŻENIE Forma CENA DATA I MIEJSCE NIERUCHOMOŚCI rozdysponow RODZAJ I w tym wywoławcza GODZINA PRZEPROWADZENIA RODZAJ ATRYBUTY l.p. (powiat/gmina/ ania CHARAKTER OGÓLNA UŻYTKI lub wywoławczy rozpoczęcia PRZETARGU PRZETARGU NIERUCHOMOŚCI obręb/numery (sprzedaż/ NIERUCHOMOŚCI /ha/ ROLNE CZYNSZ przetargu (nazwa jednostki) działek) dzierżawa /ha/ DZIERŻAWNY Powiat gryfiński 01.04.2021 Sekcja Zamiejscowa rolna niezabudowana gmina Moryń ustny 1 09.00 w Pyrzycach sprzedaż grunty orne 0,3800 0,3800 9 120,00 zł obr. Przyjezierze II nieograniczony ul. Warszawska 95 kl. IIIb, RV dz. 382 74-200 Pyrzyce Powiat gryfiński 01.04.2021 Sekcja Zamiejscowa gruntowa niezabudowana gmina Moryń ustny 2 09.30 w Pyrzycach sprzedaż łąki trwałe 0,6785 0,6785 144 480,00 zł obr. -

Integriertes Stadtentwicklungskonzept 2015 Stadt Bergen Auf Rügen
INTEGRIERTES STADTENTWICKLUNGSKONZEPT 2015 STADT BERGEN AUF RÜGEN Das vorliegende Integrierte Stadtentwicklungskonzept 2015 der Stadt Bergen a. R. wurde im Rahmen des Konzeptaufrufs des Ministeriums für Wirtschaft, Bau und Tourismus des Landes Mecklenburg- Vorpommerns für die Förderung von Projekten in der Prioritätenachse 4 „Förderung der integrierten nachhaltigen Stadtentwicklung“ erstellt. BESCHLÜSSE Beschluss der Stadtvertretung der Stadt Bergen auf Rügen am 02. Dezember 2015 IMPRESSUM Stadt Bergen a. R. Bauamt Markt 5-6 18528 Bergen auf Rügen erstellt von slapa & die raumplaner gmbh Alt-Moabit 62 10555 Berlin Tel 030 375 927 21 Fax 030 375 927 22 www.die-raumplaner.de Bearbeiter Dipl.-Ing. Sabine Slapa (strategische Leitung) Dipl.-Ing. Stefanie Burgstaller (Projektleitung) M.Sc. Niklas Steinert (Projektmitarbeit) Jana Wasicki (Projektmitarbeit) Bergen auf Rügen, 02.Dezember 2015 SEITE 2 INHALT 1 Einleitung .................................................................................................................................. 9 1.1 Anlass und Rahmenbedingungen ............................................................................................... 9 1.1 Aufgabe und Zielsetzung des ISEK 2015 ................................................................................... 10 1.2 Vorgehensweise – 2. Fortschreibung ....................................................................................... 11 2 Bestandsanalyse und Bewertung............................................................................................. -

Wykaz Identyfikatorów I Nazw Jednostek Podziału Terytorialnego Kraju” Zawiera Jednostki Tego Podziału Określone W: − Ustawie Z Dnia 24 Lipca 1998 R
ZAK£AD WYDAWNICTW STATYSTYCZNYCH, 00-925 WARSZAWA, AL. NIEPODLEG£0ŒCI 208 Informacje w sprawach sprzeda¿y publikacji – tel.: (0 22) 608 32 10, 608 38 10 PRZEDMOWA Niniejsza publikacja „Wykaz identyfikatorów i nazw jednostek podziału terytorialnego kraju” zawiera jednostki tego podziału określone w: − ustawie z dnia 24 lipca 1998 r. o wprowadzeniu zasadniczego trójstopniowego podziału terytorialnego państwa (Dz. U. Nr 96, poz. 603 i Nr 104, poz. 656), − rozporządzeniu Rady Ministrów z dnia 7 sierpnia 1998 r. w sprawie utworzenia powiatów (Dz. U. Nr 103, poz. 652) zaktualizowane na dzień 1 stycznia 2010 r. Aktualizacja ta uwzględnia zmiany w podziale teryto- rialnym kraju dokonane na podstawie rozporządzeń Rady Ministrów w okresie od 02.01.1999 r. do 01.01.2010 r. W „Wykazie...”, jako odrębne pozycje wchodzące w skład jednostek zasadniczego podziału terytorialnego kraju ujęto dzielnice m. st. Warszawy oraz delegatury (dawne dzielnice) miast: Kraków, Łódź, Poznań i Wrocław a także miasta i obszary wiejskie wchodzące w skład gmin miejsko-wiejskich. Zamieszczone w wykazie identyfikatory jednostek podziału terytorialnego zostały okre- ślone w: − załączniku nr 1 do rozporządzenia Rady Ministrów z dnia 15 grudnia 1998 r. w sprawie szczegółowych zasad prowadzenia, stosowania i udostępniania krajowego rejestru urzędo- wego podziału terytorialnego kraju oraz związanych z tym obowiązków organów admini- stracji rządowej i jednostek samorządu terytorialnego, obowiązującego od dnia 1 stycz- nia 1999 r. (Dz. U. z 1998 r. Nr 157, poz. 1031), − kolejnych rozporządzeniach Rady Ministrów zmieniających powyższe rozporządzenie w zakresie załącznika nr 1 (Dz. U. z 2000 Nr 13, poz. 161, z 2001 r. Nr 12, poz. 100 i Nr 157, poz. -

Apotropaic Practices and the Undead: a Biogeochemical Assessment of Deviant Burials in Post-Medieval Poland
RESEARCH ARTICLE Apotropaic Practices and the Undead: A Biogeochemical Assessment of Deviant Burials in Post-Medieval Poland Lesley A. Gregoricka1*, Tracy K. Betsinger2, Amy B. Scott3, Marek Polcyn4 1. Department of Sociology, Anthropology, & Social Work, University of South Alabama, Mobile, Alabama, United States of America, 2. Department of Anthropology, SUNY College at Oneonta, Oneonta, New York, United States of America, 3. Department of Anthropology, University of Manitoba, Winnipeg, Manitoba, Canada, 4. Department of Anthropology, Lakehead University, Thunder Bay, Ontario, Canada *[email protected] Abstract Apotropaic observances-traditional practices intended to prevent evil-were not OPEN ACCESS uncommon in post-medieval Poland, and included specific treatment of the dead for Citation: Gregoricka LA, Betsinger TK, Scott AB, Polcyn M (2014) Apotropaic Practices and the those considered at risk for becoming vampires. Excavations at the Drawsko 1 Undead: A Biogeochemical Assessment of Deviant cemetery (17th–18th c. AD) have revealed multiple examples (n56) of such deviant Burials in Post-Medieval Poland. PLoS ONE 9(11): e113564. doi:10.1371/journal.pone.0113564 burials amidst hundreds of normative interments. While historic records describe Editor: Sharon N. DeWitte, University of South the many potential reasons why some were more susceptible to vampirism than Carolina, United States of America others, no study has attempted to discern differences in social identity between Received: September 5, 2014 individuals within standard and deviant burials using biogeochemical analyses of Accepted: October 28, 2014 human skeletal remains. The hypothesis that the individuals selected for apotropaic Published: November 26, 2014 burial rites were non-local immigrants whose geographic origins differed from the Copyright: ß 2014 Gregoricka et al. -

West Pomerania)
zachodniopomorskie (west pomerania) Total area of the 22 892 km2 voivodeship Number of citizens: 1715,4 thous. - in urban areas 68,9 % Population per 1 km2 75 persons/km2 Principal city Szczecin Bigger cities in the Koszalin, Stargard, Kołobrzeg, Świnoujście, voivodeship Szczecinek Special Economic Zones Kostrzyńsko-Słubicka Słupska Pomorska Euro-Park Milelec Industrial & Technology Szczecin Science & Technology Park Parks Szczecin Industrial Park Goleniów Industrial Park Stargard Industrial Park High Technologies Industrial Park in Stargard Szczeciński Koszalin Technical University Industrial Park Koszalin Business Activity Zone Regional Park in Gryfino Białogard Investment Park “Invest-Park” Regional Investor Szczecin Assistance Centre International airport Goleniów West Pomerania’s economy depends on its location - direct access to the Baltic Sea (through the port of Szczecin), and proximity to Germany and the Scandinavian countries. Distance between Szczecin - European seaport and the capital of the province, and Berlin is only 130 km. The region, primarily associated with the shipbuilding industry and agriculture, today develops various sectors of the economy, especially those related to advanced technologies. Region as more and more uses renewable energy sources, which can be observed by increasing the number of wind turbines and thermal power plants. Investment opportunities • direct access to the sea • situated at the junction of important international transport routes (North-South and West-East) • sited alongside the inside -

Official Journal L 329 of the European Union
Official Journal L 329 of the European Union ★ ★ ★ ★ ★ ★ ★ ★ ★ ★ ★ ★ Volume 59 English edition Legislation 3 December 2016 Contents II Non-legislative acts INTERNATIONAL AGREEMENTS ★ Council Decision (Euratom) 2016/2116 of 12 February 2016 approving the conclusion by the European Commission, on behalf of the European Atomic Energy Community, of the Agreement extending the Framework Agreement for International Collaboration on Research and Development of Generation IV nuclearenergy systems .................................................. 1 Agreement extending the Framework Agreement for International Collaboration on Research and Development of Generation IV nuclearenergy systems ...................................................................... 3 ★ Council Decision (EU) 2016/2117 of 29 September 2016 on the conclusion, on behalf of the Union, of the Framework Agreement on Comprehensive Partnership and Cooperation between the European Union and its Member States, of the one part, and the Socialist Republic of Vietnam, of the other part .............................................................................. 6 Framework Agreement on Comprehensive Partnership and Cooperation between the European Union and its Member States, of the one part, and the Socialist Republic of Viet Nam, of the other part ................... 8 ★ Council Decision (EU) 2016/2118 of 28 October 2016 on the signing, on behalf of the Union, and provisional application of the Strategic Partnership Agreement between the European Union and its Member States, -
Available As PDF
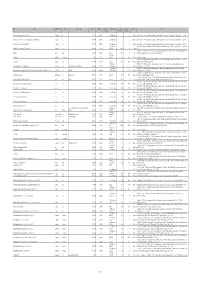
ID Locality Administrative unit Country Alternate spelling Latitude Longitude Geographic Stratigraphic age Age, lower Age, upper Epoch Reference precision boundary boundary 3247 Acquasparta (along road to Massa Martana) Acquasparta Italy 42.707778 12.552333 1 late Villafranchian 2 1.1 Pleistocene Esu, D., Girotti, O. 1975. La malacofauna continentale del Plio-Pleistocene dell’Italia centrale. I. Paleontologia. Geologica Romana, 13, 203-294. 3246 Acquasparta (NE of 'km 32', below Chiesa di Santa Lucia di Burchiano) Acquasparta Italy 42.707778 12.552333 1 late Villafranchian 2 1.1 Pleistocene Esu, D., Girotti, O. 1975. La malacofauna continentale del Plio-Pleistocene dell’Italia centrale. I. Paleontologia. Geologica Romana, 13, 203-294. 3234 Acquasparta (Via Tiberina, 'km 32,700') Acquasparta Italy 42.710556 12.549556 1 late Villafranchian 2 1.1 Pleistocene Esu, D., Girotti, O. 1975. La malacofauna continentale del Plio-Pleistocene dell’Italia centrale. I. Paleontologia. Geologica Romana, 13, 203-294. upper Alluvial Ewald, R. 1920. Die fauna des kalksinters von Adelsheim. Jahresberichte und Mitteilungen des Oberrheinischen geologischen Vereines, Neue Folge, 3330 Adelsheim (Adelsheim, eastern part of the city) Adelsheim Germany 49.402305 9.401456 2 (Holocene) 0.00585 0 Holocene 9, 15-17. Sanko, A.F. 2007. Quaternary freshwater mollusks Belarus and neighboring regions of Russia, Lithuania, Poland (field guide). [in Russian]. Institute 5200 Adrov Adrov Belarus 54.465816 30.389993 2 Holocene 0.0117 0 Holocene of Geochemistry and Geophysics, National Academy of Sciences, Belarus. middle-late 4441 Adzhikui Adzhikui Turkmenistan 39.76667 54.98333 2 Pleistocene 0.781 0.0117 Pleistocene FreshGEN team decision Hagemann, J. 1976. Stratigraphy and sedimentary history of the Upper Cenozoic of the Pyrgos area (Western Peloponnesus), Greece. -

Kindertagesstätten Der Volkssolidarität (389) Stand September 2020
Kindertagesstätten der Volkssolidarität (389) Stand September 2020 Name / Kontaktdaten Rahmenbedingungen Besonderheiten der pädagogischen Arbeit Landesverband Berlin (8 Kitas) Kita gGmbH Geschäftsführerin: Dr. Ute Gerwert, 030 / 403 66 20 - 00, [email protected], [email protected] Fachberatung & QM-Beauftragte: Daniela Wiegert, 030 / 402 66 20 – 30, [email protected] Kinderschutz, Koordination Familienzentrumsarbeit: Krümmel, Elke, 030 / 403 66 20 – 31, [email protected] Familienzentrumsarbeit in allen Kitas nach Bedarf regelmäßige Übernachtungen mobile Beratung des Kinder- und Jugend-Gesundheitsdienstes (KJGD) ABC-Schülertreff Familienfreizeit nach §16 Abs. 3 Elternkurse „Starke Eltern – Starke Kinder“ Jugendberatung Kita „Regenbogeninsel“ 240 Plätze Familienergänzend, integrativ, interkulturell Leitung: Bärbel Schröder, Ina Schultgen ab 8. Wo. bis zum Schuleintritt Sauna Glambecker Ring 50/52 Betreuung/Förderung von Integra- selbstkochende Einrichtung 12679 Berlin tionskindern Angebote in Form von Familienzentrumsarbeit (z.B. Krabbelgruppen, Elterncafés, Über- Telefon: (030)9328119 6:00 – 18:00 Uhr nachtung, Vater-Kind-Angebote, Familienfreizeitangebote am Wochenende) kita-regenbogeninsel-marzahn@volkssoli- bei Bedarf bis 19:30 Uhr daritaet.de Offene Arbeit (etagenübergrei- fend) Arbeit nach dem Situationsansatz Kita „Kleine Traber“ 179 Plätze familienergänzend, integrativ und interkulturell. Treskowallee 135 ab 8. Wo. bis zum Schuleintritt Altersmischung -

DOKUMENTATION Der Städtepartnerschaftlichen Und Internationalen Kontakte
DOKUMENTATION der städtepartnerschaftlichen und internationalen Kontakte STADT BErgEN AUf rügEN in den Jahren 1990 – 2009 DOKUMENTATION der städtepartnerschaftlichen und internationalen Kontakte der Stadt BErgen auf rügen in den Jahren 1990 – 2009 DOKUMENTATION Vorwort Sehr geehrte Damen und Herren! Die Ihnen vorliegende Dokumentation beinhaltet eindrucksvoll die städ- tepartnerschaftlichen Beziehungen und die internationalen Aktivitäten der Stadt Bergen auf Rügen. Neben den seit 1990 möglichen innerdeutschen Kontakten entstanden aufrichtige Freundschaften über die Ostsee nach Skandinavien und zum polnischen Nachbarland. So besteht seit 20 Jahren eine intensive Partnerschaft zur Stadt Oldenburg in Holstein und später eine im Drei- erverbund funktionierende Freundschaft mit der schwedischen Stadt Svedala und der polnischen Stadt Goleniow. Diese Kontakte werden nicht nur von den politisch Verantwortlichen gepflegt, sondern waren und sind vor allem Begegnungen unserer Bürgerinnen und Bürger, unserer Kinder und Jugendlichen und der Mitglieder von Vereinen aller gesellschaftlicher Bereiche. Darüber hinaus ist die Stadt Bergen auf Rügen durch ihre territoriale Lage und ihre Stellung als Mit- telzentrum der Insel Rügen oft Austragungsort überregionaler, nationaler und europäischer Veranstal- tungen und ist dabei immer an weiteren internationalen Kontakten auf wirtschaftlichen oder kulturellen Gebieten interessiert. Bergen auf Rügen profiliert sich aber auch als Stadt der Zukunft für Bildung und Gesundheit. Dafür wurden bereits wichtige Voraussetzungen