Wallace and the Species Concept of the Early Darwinians
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Social Mimesis As an Argument in Evolutionary Theory Around 1900
Hist. Phil. Life Sci., 31 (2009), 201-214 “Imitation of Similar Beings”: Social Mimesis as an Argument in Evolutionary Theory around 1900 Stefan Willer Zentrum für Literatur- und Kulturforschung Schützenstraße 18 10117 Berlin, Germany ABSTRACT – The article analyzes imitation as both a fascinating and irritating phenomenon in “classical” evolutionary theory. Evolutionists situate imitation on the threshold between the natural and the socio-cultural, hence between the animal and the human. This intermediate position can be regarded as a symptom for the unresolved and maybe unresolvable problem of intentionality and teleology in nature. To elaborate this problem, I examine the ways in which imitation was conceived of by the German Africologist Wilhelm Bleek in his treatise On the Origin of Language and by Charles Darwin in The Descent of Man. Bleek and Darwin share a high esteem of imitation, which they see as the mainspring of human mental capacities, including language. But at the same time, imitation for them is the epitome of a low level of consciousness, embodied in the figures of the idiot, the savage, and the ape. Thus, the problem of similarity comes to the fore: similarity produced by imitation, but also being at the basis of every act of imitation. This problem is further evidenced with a side glance on Darwin’s remarks about mimicry in The Origin of Species. The article closes with a literary reading of Franz Kafka’s Report to an Academy, in which imitation and similarity represent survival strategies and motivate a strange shift from ape to man. KEYWORDS – Ape, imitation, intention, language, mimesis, mimicry, resemblance, similarity, survival Introduction To begin, the current relevance of the problem of imitation needs to be stressed. -

A Database Integrating Genomic Indel Polymorphisms That Distinguish Mouse Strains Keiko Akagi1,2, Robert M
D600–D606 Nucleic Acids Research, 2010, Vol. 38, Database issue Published online 20 November 2009 doi:10.1093/nar/gkp1046 MouseIndelDB: a database integrating genomic indel polymorphisms that distinguish mouse strains Keiko Akagi1,2, Robert M. Stephens3, Jingfeng Li2,4, Evgenji Evdokimov5, Michael R. Kuehn5, Natalia Volfovsky3 and David E. Symer2,4,6,7,8,9,* 1Mouse Cancer Genetics Program, Center for Cancer Research, National Cancer Institute, Frederick, MD 21702, 2Department of Molecular Virology, Immunology and Medical Genetics, The Ohio State University Comprehensive Cancer Center, Columbus, OH 43210, 3Advanced Biomedical Computing Center, Information Systems Program, SAIC-Frederick, Inc., NCI-Frederick, Frederick, MD 21702, 4Basic Research Laboratory, 5Laboratory of Protein Dynamics and Signaling, 6Laboratory of Biochemistry and Molecular Biology, Center for Cancer Research, National Cancer Institute, Frederick, MD 21702, 7Human Cancer Genetics Program, 8Departments of Internal Medicine and 9Biomedical Informatics, The Ohio State University Comprehensive Cancer Center, Columbus, OH 43210, USA Received August 5, 2009; Revised October 23, 2009; Accepted October 27, 2009 ABSTRACT wide-ranging functional differences. This extensive phenotypic variation has helped to make the mouse a MouseIndelDB is an integrated database resource premier model organism, mimicking many aspects of containing thousands of previously unreported human diversity and diseases. Understanding the mouse genomic indel (insertion and deletion) genomic differences that -

Pathway in Protective Immunity Jeremy Manry, Guillaume Laval, Etienne Patin, Simona Fornarino, Christiane Bouchier, Magali Tichit, Luis Barreiro, Lluis Quintana-Murci
Evolutionary genetics evidence of an essential, non-redundant role of the IFN-γ pathway in protective immunity Jeremy Manry, Guillaume Laval, Etienne Patin, Simona Fornarino, Christiane Bouchier, Magali Tichit, Luis Barreiro, Lluis Quintana-Murci To cite this version: Jeremy Manry, Guillaume Laval, Etienne Patin, Simona Fornarino, Christiane Bouchier, et al.. Evo- lutionary genetics evidence of an essential, non-redundant role of the IFN-γ pathway in protective immunity. Human Mutation, Wiley, 2011, 32 (6), pp.633-42. 10.1002/humu.21484. hal-00627541 HAL Id: hal-00627541 https://hal.archives-ouvertes.fr/hal-00627541 Submitted on 29 Sep 2011 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Human Mutation Evolutionary genetics evidence of an essential, non- redundant role of the IFN-γ pathway in protective immunity For Peer Review Journal: Human Mutation Manuscript ID: humu-2010-0462.R1 Wiley - Manuscript type: Research Article Date Submitted by the 23-Jan-2011 Author: Complete List of Authors: Manry, Jeremy; Institut Pasteur Laval, Guillaume; Institut Pasteur Patin, Etienne; Institut Pasteur Fornarino, Simona; Institut Pasteur Bouchier, Christiane; Institut Pasteur Tichit, Magali; Institut Pasteur Barreiro, Luis; University of Chicago Quintana-Murci, Lluis; Institut Pasteur, EEMI IFNG, IFNGR1, IFNGR2, natural selection, polymorphism, Key Words: population genetics John Wiley & Sons, Inc. -

Alfred Russel Wallace and the Darwinian Species Concept
Gayana 73(2): Suplemento, 2009 ISSN 0717-652X ALFRED RUSSEL WALLACE AND THE Darwinian SPECIES CONCEPT: HIS paper ON THE swallowtail BUTTERFLIES (PAPILIONIDAE) OF 1865 ALFRED RUSSEL WALLACE Y EL concepto darwiniano DE ESPECIE: SU TRABAJO DE 1865 SOBRE MARIPOSAS papilio (PAPILIONIDAE) Jam ES MA LLET 1 Galton Laboratory, Department of Biology, University College London, 4 Stephenson Way, London UK, NW1 2HE E-mail: [email protected] Abstract Soon after his return from the Malay Archipelago, Alfred Russel Wallace published one of his most significant papers. The paper used butterflies of the family Papilionidae as a model system for testing evolutionary hypotheses, and included a revision of the Papilionidae of the region, as well as the description of some 20 new species. Wallace argued that the Papilionidae were the most advanced butterflies, against some of his colleagues such as Bates and Trimen who had claimed that the Nymphalidae were more advanced because of their possession of vestigial forelegs. In a very important section, Wallace laid out what is perhaps the clearest Darwinist definition of the differences between species, geographic subspecies, and local ‘varieties.’ He also discussed the relationship of these taxonomic categories to what is now termed ‘reproductive isolation.’ While accepting reproductive isolation as a cause of species, he rejected it as a definition. Instead, species were recognized as forms that overlap spatially and lack intermediates. However, this morphological distinctness argument breaks down for discrete polymorphisms, and Wallace clearly emphasised the conspecificity of non-mimetic males and female Batesian mimetic morphs in Papilio polytes, and also in P. -

Naturalist Narratives: the Literary Tradition Among Victorian Explorers of the Amazon
NATURALIST NARRATIVES: THE LITERARY TRADITION AMONG VICTORIAN EXPLORERS OF THE AMAZON Frank Izaguirre1 Abstract: Victorian explorers of the Amazon are remembered the world over for their vast and varied contributions to science: from Henry Walter Bates’s observations of mimicry in animals to Alfred Russell Wallace’s co-discovery of natural selection. Yet an under-examined and essential element of the lives of these explorer-naturalists was their ethic of writing, how they catalogued their adventures in narratives that not only captivated the world, but influenced the development of scientific exploration in the region and led to the birth of a unique subgenre of environmental writing. In the study, I will show how the natural narratives, principally Henry Walter Bates’s The Naturalist on the River Amazons and Alfred Russell Wallace’s A Narrative of Travels on the Amazon and Rio Negro: With an Account of the Native Tribes influenced the decisions future naturalists would make about their own journeys, thereby shaping the advancement of scientific understanding of the Amazon as well as the course of its exploration by the western world. Keywords: Amazon. Naturalist. Edwards. Bates. Wallace. Resumo: Exploradores vitorianos da Amazônia são lembrados em todo o mundo por suas vastas e variadas contribuições para a Ciência, a partir de observações sobre o mimetismo animal, desde Henry Walter Bates até a seleção natural de Alfred Russell Wallace. Um dos elementos essenciais ainda não completamente examinados até hoje são seus escritos éticos e como eles catalogaram suas aventuras em narrativas que não só cativaram o mundo, como também influenciaram o desenvolvimento da exploração científica na região que levou ao nascimento de um subgênero da escrita ambiental. -
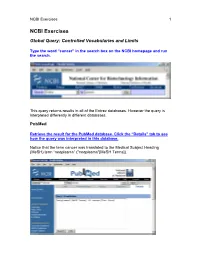
NCBI Exercises 1
NCBI Exercises 1 NCBI Exercises Global Query: Controlled Vocabularies and Limits Type the word “cancer” in the search box on the NCBI homepage and run the search. This query returns results in all of the Entrez databases. However the query is interpreted differently in different databases. PubMed Retrieve the result for the PubMed database. Click the “Details” tab to see how the query was interpreted in this database. Notice that the term cancer was translated to the Medical Subject Heading (MeSH) term “neoplasms” ("neoplasms"[MeSH Terms]). NCBI Exercises 2 MeSH is a controlled vocabulary that is used to index all articles in PubMed. In the details box, edit the query to remove the portion that searched for cancer as a text word and run the search. Notice that the number of articles retrieved has changed. These will be a more relevant set of results. You can force the PubMed engine to only search the MeSH vocabulary or specify any other indexed field through the “Limits” tab. Use the Web browser’s back button to return to the Global query page and retrieve the PubMed results again. Now click on the “Limits” tab. Select “MeSH terms” from the first drop-down menu, the one headed by “All Fields”. Now run the search with the limit in place and check the “Details” tab to verify that only the MeSH term translation was used. Nucleotide Use the Web browser’s back button to return to the Global query page. Retrieve the results for the nucleotide database. Click the “Details” tab to see how the query was interpreted for this molecular database. -

Natural Selection: Charles Darwin & Alfred Russel Wallace
Search | Glossary | Home << previous | next > > Natural Selection: Charles Darwin & Alfred Russel Wallace The genius of Darwin (left), the way in which he suddenly turned all of biology upside down in 1859 with the publication of the Origin of Species , can sometimes give the misleading impression that the theory of evolution sprang from his forehead fully formed without any precedent in scientific history. But as earlier chapters in this history have shown, the raw material for Darwin's theory had been known for decades. Geologists and paleontologists had made a compelling case that life had been on Earth for a long time, that it had changed over that time, and that many species had become extinct. At the same time, embryologists and other naturalists studying living animals in the early 1800s had discovered, sometimes unwittingly, much of the A visit to the Galapagos Islands in 1835 helped Darwin best evidence for Darwin's formulate his ideas on natural selection. He found theory. several species of finch adapted to different environmental niches. The finches also differed in beak shape, food source, and how food was captured. Pre-Darwinian ideas about evolution It was Darwin's genius both to show how all this evidence favored the evolution of species from a common ancestor and to offer a plausible mechanism by which life might evolve. Lamarck and others had promoted evolutionary theories, but in order to explain just how life changed, they depended on speculation. Typically, they claimed that evolution was guided by some long-term trend. Lamarck, for example, thought that life strove over time to rise from simple single-celled forms to complex ones. -

The Genus Acraea (Lepidoptera : Nymphalidae) - Peter Hendry
The genus Acraea (Lepidoptera : Nymphalidae) - Peter Hendry With the recent migration to Australia of the Tawny Coster (Acraea terpsicore (Linnaeus, 1758)), (see Creature Feature this issue), I thought it might be timely to take a look at the genus worldwide. It must be noted that due to a misidentification A. terpsicore had long been known as A. violae and many references in the literature and on the web refer to it as A. violae. As with much of the Lepidoptera the genus is in a state of flux, and has long been split into the subgenera Acraea (Acraea) and Acraea (Actinote). The genus is placed in the tribe Acraeini and until Harvey (1991) placed it in the subfamily Heliconiinae it was listed in the subfamily Acraeinae. Recent molecular work has made changes and a current listing of the tribe Acraeini, by Niklas Wahlberg, is available at http://www.nymphalidae.net/Classification/Acraeini.htm. It shows members of the old subgenus Acraea (Actinote) being placed in the genus Actinote, and the old subgenus Acraea (Acraea) becoming the genus Acraea with a subgenus Acraea (Bematistes). It also lists several Acraea as unplaced. This may further change as some believe the subgenus Acraea (Bematistes) will move to the genus Bematistes. The genus is primarily Afrotropical with only four species occurring outside this region, these being, Acraea andromacha (Fig. 1) A. meyeri (Fig. 10) A. moluccana and A. terpsicore. A fifth species the Yellow Coster Acraea (Actinote) issoria is now referred to the genus Actinote. Like many of the Nymphalidae the larvae feed on plants which contain cyanogens making the larvae and adults poisonous to predators. -

The Speciation History of Heliconius: Inferences from Multilocus DNA Sequence Data
The speciation history of Heliconius: inferences from multilocus DNA sequence data by Margarita Sofia Beltrán A thesis submitted for the degree of Doctor of Philosophy of the University of London September 2004 Department of Biology University College London 1 Abstract Heliconius butterflies, which contain many intermediate stages between local varieties, geographic races, and sympatric species, provide an excellent biological model to study evolution at the species boundary. Heliconius butterflies are warningly coloured and mimetic, and it has been shown that these traits can act as a form of reproductive isolation. I present a species-level phylogeny for this group based on 3834bp of mtDNA (COI, COII, 16S) and nuclear loci (Ef1α, dpp, ap, wg). Using these data I test the geographic mode of speciation in Heliconius and whether mimicry could drive speciation. I found little evidence for allopatric speciation. There are frequent shifts in colour pattern within and between sister species which have a positive and significant correlation with species diversity; this suggests that speciation is facilitated by the evolution of novel mimetic patterns. My data is also consistent with the idea that two major innovations in Heliconius, adult pollen feeding and pupal-mating, each evolved only once. By comparing gene genealogies from mtDNA and introns from nuclear Tpi and Mpi genes, I investigate recent speciation in two sister species pairs, H. erato/H. himera and H. melpomene/H. cydno. There is highly significant discordance between genealogies of the three loci, which suggests recent speciation with ongoing gene flow. Finally, I explore the phylogenetic relationships between races of H. melpomene using an AFLP band tightly linked to the Yb colour pattern locus (which determines the yellow bar in the hindwing). -

Lepidoptera: Nymphalidae) En Dos Especies De Passiflora
R158evista Colombiana de Entomología 36 (1): 158-164 (2010) Desarrollo, longevidad y oviposición de Heliconius charithonia (Lepidoptera: Nymphalidae) en dos especies de Passiflora Development, longevity, and oviposition of Heliconius charithonia (Lepidoptera: Nymphalidae) on two species of Passiflora CAROLINA MILLÁN J.1, PATRICIA CHACÓN C.2 y GERMÁN CORREDOR3 Resumen: El desarrollo de Heliconius charithonia en dos especies de plantas hospederas se estudió en el mariposario del Zoológico de Cali (Colombia) entre diciembre de 2007 y octubre de 2008. Se siguió el desarrollo de larvas pro- venientes de 90 y 83 huevos puestos en Passiflora adenopoda y P. rubra respectivamente. Se midió la duración de los cinco instares larvales, así como el peso y longitud pupal. Los adultos emergidos se marcaron, midieron, sexaron y se liberaron en el área de exhibición del mariposario y se hicieron censos semanales para estimar la longevidad. La sobrevivencia larval fue mayor en P. adenopoda (76,4%) con respecto a P. rubra (33,9%). La mortalidad pupal alcanzó un 3% en P. rubra mientras en P. adenopoda todas las pupas fueron viables. Los resultados idican que P. adenopoda es el hospedero de oviposición más propicio para la cría masiva de H. charithonia, ya que en dicho hospedero se observó un mejor desarrollo larval, pupas más grandes y más pesadas, y los adultos mostraron mayor longitud alar y mayor longevidad (140 días vs 70 días). La preferencia de oviposición mostraron que del total de huevos (N = 357) el 71% fué depositado sobre P. adenopoda, aun en aquellos casos en que las hembras se desarrollaron sobre P. rubra. -

The History and Influence of Maria Sibylla Merian's Bird-Eating Tarantula: Circulating Images and the Production of Natural Knowledge
Biology Faculty Publications Biology 2016 The History and Influence of Maria Sibylla Merian's Bird-Eating Tarantula: Circulating Images and the Production of Natural Knowledge Kay Etheridge Gettysburg College Follow this and additional works at: https://cupola.gettysburg.edu/biofac Part of the Biology Commons, and the Illustration Commons Share feedback about the accessibility of this item. Recommended Citation Etheridge, K. "The History and Influence of Maria Sibylla Merian’s Bird-Eating Tarantula: Circulating Images and the Production of Natural Knowledge." Global Scientific Practice in the Age of Revolutions, 1750 – 1850. P. Manning and D. Rood, eds. (Pittsburgh, University of Pittsburgh Press. 2016). 54-70. This is the publisher's version of the work. This publication appears in Gettysburg College's institutional repository by permission of the copyright owner for personal use, not for redistribution. Cupola permanent link: https://cupola.gettysburg.edu/biofac/54 This open access book chapter is brought to you by The Cupola: Scholarship at Gettysburg College. It has been accepted for inclusion by an authorized administrator of The Cupola. For more information, please contact [email protected]. The History and Influence of Maria Sibylla Merian's Bird-Eating Tarantula: Circulating Images and the Production of Natural Knowledge Abstract Chapter Summary: A 2009 exhibition at the Fitzwilliam Museum on the confluence of science and the visual arts included a plate from a nineteenth-century encyclopedia owned by Charles Darwin showing a tarantula poised over a dead bird (figure 3.1).1 The genesis of this startling scene was a work by Maria Sibylla Merian (German, 1647–1717), and the history of this image says much about how knowledge of the New World was obtained, and how it was transmitted to the studies and private libraries of Europe, and from there into popular works like Darwin’s encyclopedia. -

The Malay Archipelago
BOOKS & ARTS COMMENT The Malay Archipelago: the land of the orang-utan, and the bird of paradise; a IN RETROSPECT narrative of travel, with studies of man and nature ALFRED RUSSEL WALLACE The Malay Macmillan/Harper Brothers: first published 1869. lfred Russel Wallace was arguably the greatest field biologist of the nine- Archipelago teenth century. He played a leading Apart in the founding of both evolutionary theory and biogeography (see page 162). David Quammen re-enters the ‘Milky Way of He was also, at times, a fine writer. The best land masses’ evoked by Alfred Russel Wallace’s of his literary side is on show in his 1869 classic, The Malay Archipelago, a wondrous masterpiece of biogeography. book of travel and adventure that wears its deeper significance lightly. The Malay Archipelago is the vast chain of islands stretching eastward from Sumatra for more than 6,000 kilometres. Most of it now falls within the sovereignties of Malaysia and Indonesia. In Wallace’s time, it was a world apart, a great Milky Way of land masses and seas and straits, little explored by Europeans, sparsely populated by peoples of diverse cul- tures, and harbouring countless species of unknown plant and animal in dense tropical forests. Some parts, such as the Aru group “Wallace paid of islands, just off the his expenses coast of New Guinea, by selling ERNST MAYR LIB., MUS. COMPARATIVE ZOOLOGY, HARVARD UNIV. HARVARD ZOOLOGY, LIB., MUS. COMPARATIVE MAYR ERNST were almost legend- specimens. So ary for their remote- he collected ness and biological series, not just riches. Wallace’s jour- samples.” neys throughout this region, sometimes by mail packet ship, some- times in a trading vessel or a small outrigger canoe, were driven by a purpose: to collect animal specimens that might help to answer a scientific question.