Development of a Functional in Vitro 3D Model of the Peripheral Nerve
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
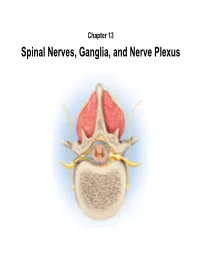
Spinal Nerves, Ganglia, and Nerve Plexus Spinal Nerves
Chapter 13 Spinal Nerves, Ganglia, and Nerve Plexus Spinal Nerves Posterior Spinous process of vertebra Posterior root Deep muscles of back Posterior ramus Spinal cord Transverse process of vertebra Posterior root ganglion Spinal nerve Anterior ramus Meningeal branch Communicating rami Anterior root Vertebral body Sympathetic ganglion Anterior General Anatomy of Nerves and Ganglia • Spinal cord communicates with the rest of the body by way of spinal nerves • nerve = a cordlike organ composed of numerous nerve fibers (axons) bound together by connective tissue – mixed nerves contain both afferent (sensory) and efferent (motor) fibers – composed of thousands of fibers carrying currents in opposite directions Anatomy of a Nerve Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Epineurium Perineurium Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Endoneurium Nerve Rootlets fiber Posterior root Fascicle Posterior root ganglion Anterior Blood root vessels Spinal nerve (b) Copyright by R.G. Kessel and R.H. Kardon, Tissues and Organs: A Text-Atlas of Scanning Electron Microscopy, 1979, W.H. Freeman, All rights reserved Blood vessels Fascicle Epineurium Perineurium Unmyelinated nerve fibers Myelinated nerve fibers (a) Endoneurium Myelin General Anatomy of Nerves and Ganglia • nerves of peripheral nervous system are ensheathed in Schwann cells – forms neurilemma and often a myelin sheath around the axon – external to neurilemma, each fiber is surrounded by -

The Peripheral Nervous System
The Peripheral Nervous System Dr. Ali Ebneshahidi Peripheral Nervous System (PNS) – Consists of 12 pairs of cranial nerves and 31 pairs of spinal nerves. – Serves as a critical link between the body and the central nervous system. – peripheral nerves contain an outermost layer of fibrous connective tissue called epineurium which surrounds a thinner layer of fibrous connective tissue called perineurium (surrounds the bundles of nerve or fascicles). Individual nerve fibers within the nerve are surrounded by loose connective tissue called endoneurium. Cranial Nerves Cranial nerves are direct extensions of the brain. Only Nerve I (olfactory) originates from the cerebrum, the remaining 11 pairs originate from the brain stem. Nerve I (Olfactory)- for the sense of smell (sensory). Nerve II (Optic)- for the sense of vision (sensory). Nerve III (Oculomotor)- for controlling muscles and accessory structures of the eyes ( primarily motor). Nerve IV (Trochlear)- for controlling muscles of the eyes (primarily motor). Nerve V (Trigeminal)- for controlling muscles of the eyes, upper and lower jaws and tear glands (mixed). Nerve VI (Abducens)- for controlling muscles that move the eye (primarily motor). Nerve VII (Facial) – for the sense of taste and controlling facial muscles, tear glands and salivary glands (mixed). Nerve VIII (Vestibulocochlear)- for the senses of hearing and equilibrium (sensory). Nerve IX (Glossopharyngeal)- for controlling muscles in the pharynx and to control salivary glands (mixed). Nerve X (Vagus)- for controlling muscles used in speech, swallowing, and the digestive tract, and controls cardiac and smooth muscles (mixed). Nerve XI (Accessory)- for controlling muscles of soft palate, pharynx and larynx (primarily motor). Nerve XII (Hypoglossal) for controlling muscles that move the tongue ( primarily motor). -
PN1 (Midha) Microanatomy of Peripheral Nerves-Part1.Pdf
Peripheral Nerve Basics • Consist of processes of cell bodies found in the DRG, anterior horn, and autonomic ganglia • Organized by several distinct connective tissue layers – the epineurium, perineurium, and endoneurium • Vascular supply provided by the vasa nervorum Peripheral Nerve Basics • Neuronal processes bound into fascicles by perineurium • Fascicles bound into nerves by epineurium • Endoneurium is a division of the perineurium which form thin layers of connective tissue surrounding neuronal fibers in a fascicle Sural nerve in cross-section Epineurium • Loose areolar tissue with sparse, longitudinally-oriented collagen fibers • Some elastic fibers where epineurium abuts perineurium • Able to accommodate a significant amount of nerve stretching and movement • Increases in thickness where nerves cross joints • Constitutes an increasing proportion of nerves as they increase in size • Epineurial fat helps cushion nerves from compressive injury • Decreased epineurial fat found in patients with diabetes Perineurium • Cellular component composed of laminated fibroblasts of up to 15 layers in thickness which are bounded by a basal lamina • Semi-permeable: inner lamellae have tight junctions, providing a barrier to intercellular transport of macromolecules – Tight junctions can be loosened with topical anaesthetics and with osmotic change Perineurium • Exhibits a slightly positive internal pressure – Fascicular contents herniate upon perineurial injury • Under tension longitudinally – Nerve segment shortens upon transection – may complicate surgical repair as nerve can be stretched only approximately 10% before being inhibited by collagen Endoneurium • Intrafascicular connective tissue consisting of a collagenous matrix in the interstitial space • Develops into partitions of dense connective tissue between diverging fascicles and eventually becomes perineurium when the fascicles separate • Collagen fibers are longitudinally-oriented and run along nerve fibers and capillaries. -

Modulatory Roles of ATP and Adenosine in Cholinergic Neuromuscular Transmission
International Journal of Molecular Sciences Review Modulatory Roles of ATP and Adenosine in Cholinergic Neuromuscular Transmission Ayrat U. Ziganshin 1,* , Adel E. Khairullin 2, Charles H. V. Hoyle 1 and Sergey N. Grishin 3 1 Department of Pharmacology, Kazan State Medical University, 49 Butlerov Street, 420012 Kazan, Russia; [email protected] 2 Department of Biochemistry, Laboratory and Clinical Diagnostics, Kazan State Medical University, 49 Butlerov Street, 420012 Kazan, Russia; [email protected] 3 Department of Medical and Biological Physics with Computer Science and Medical Equipment, Kazan State Medical University, 49 Butlerov Street, 420012 Kazan, Russia; [email protected] * Correspondence: [email protected]; Tel.: +7-843-236-0512 Received: 30 June 2020; Accepted: 1 September 2020; Published: 3 September 2020 Abstract: A review of the data on the modulatory action of adenosine 5’-triphosphate (ATP), the main co-transmitter with acetylcholine, and adenosine, the final ATP metabolite in the synaptic cleft, on neuromuscular transmission is presented. The effects of these endogenous modulators on pre- and post-synaptic processes are discussed. The contribution of purines to the processes of quantal and non- quantal secretion of acetylcholine into the synaptic cleft, as well as the influence of the postsynaptic effects of ATP and adenosine on the functioning of cholinergic receptors, are evaluated. As usual, the P2-receptor-mediated influence is minimal under physiological conditions, but it becomes very important in some pathophysiological situations such as hypothermia, stress, or ischemia. There are some data demonstrating the same in neuromuscular transmission. It is suggested that the role of endogenous purines is primarily to provide a safety factor for the efficiency of cholinergic neuromuscular transmission. -

Nomina Histologica Veterinaria, First Edition
NOMINA HISTOLOGICA VETERINARIA Submitted by the International Committee on Veterinary Histological Nomenclature (ICVHN) to the World Association of Veterinary Anatomists Published on the website of the World Association of Veterinary Anatomists www.wava-amav.org 2017 CONTENTS Introduction i Principles of term construction in N.H.V. iii Cytologia – Cytology 1 Textus epithelialis – Epithelial tissue 10 Textus connectivus – Connective tissue 13 Sanguis et Lympha – Blood and Lymph 17 Textus muscularis – Muscle tissue 19 Textus nervosus – Nerve tissue 20 Splanchnologia – Viscera 23 Systema digestorium – Digestive system 24 Systema respiratorium – Respiratory system 32 Systema urinarium – Urinary system 35 Organa genitalia masculina – Male genital system 38 Organa genitalia feminina – Female genital system 42 Systema endocrinum – Endocrine system 45 Systema cardiovasculare et lymphaticum [Angiologia] – Cardiovascular and lymphatic system 47 Systema nervosum – Nervous system 52 Receptores sensorii et Organa sensuum – Sensory receptors and Sense organs 58 Integumentum – Integument 64 INTRODUCTION The preparations leading to the publication of the present first edition of the Nomina Histologica Veterinaria has a long history spanning more than 50 years. Under the auspices of the World Association of Veterinary Anatomists (W.A.V.A.), the International Committee on Veterinary Anatomical Nomenclature (I.C.V.A.N.) appointed in Giessen, 1965, a Subcommittee on Histology and Embryology which started a working relation with the Subcommittee on Histology of the former International Anatomical Nomenclature Committee. In Mexico City, 1971, this Subcommittee presented a document entitled Nomina Histologica Veterinaria: A Working Draft as a basis for the continued work of the newly-appointed Subcommittee on Histological Nomenclature. This resulted in the editing of the Nomina Histologica Veterinaria: A Working Draft II (Toulouse, 1974), followed by preparations for publication of a Nomina Histologica Veterinaria. -

The Coolest Innovation in Pain Management
A NEW WAY TO FREEZE OUT Provides Temporary POST-OPERATIVE PAIN. Post-Operative Pain Relief Cryo Nerve Block (cryoNB) harnesses the power of cryotherapy to provide a new form of temporary pain relief for patients who undergo certain invasive cardiac or thoracic surgical procedures. This technology uses a unique freezing method to temporarily block pain signals from nerves in the affected area, providing an effect that’s similar to a local anesthetic. ATRICURE, INC. Indications for Use: AtriCure’s cryoICE� cryoSPHERE™ cryoablation probes are sterile, single use devices 7555 Innovation Way intended for use in blocking pain by temporarily ablating Mason, Ohio 45040 peripheral nerves. The Coolest Innovation +1 (513) 755-4100 Please review the Instructions for Use for a complete +1 (888) 347-6403 listing of contraindications, warnings, precautions and potential adverse events prior to using these devices. in Pain Management. Customer Service / Product Inquiries Rx Only. Phone: +1 (866) 349-2342 Fax: +1 (513) 755-4567 PM-US-0017A-0920-G HOW cryoNB WORKS. 2 / STOPPING THE PAIN SIGNALS A NEW APPLICATION FOR Freezing blocks pain signals, which originate from a structure within the A PROVEN TECHNOLOGY. 1 / FREEZING THE NERVE nerve called the axon. The axons subsequently degenerate while leaving the A surgical probe delivers extremely cold temperature to nerve tissue in the tubule structures (epineurium, perineurium, endoneurium) intact for the Cryotherapy was originally developed as a treatment for cardiac nerves around the surgical incision. axons to regenerate within. arrhythmias. AtriCure consulted with leading pain management experts to develop new techniques for applying cryoNB to nerves, specifically in order to block pain. -

Journal of Biomechanical Science and Engineering Researches on Biomechanical Properties and Models of Peripheral Nerves
Bulletin of the JSME Vol.12, No.1, 2017 Journal of Biomechanical Science and Engineering 【Review Paper】 Researches on biomechanical properties and models of peripheral nerves - a review Ming-Shaung JU*, Chou-Ching K. LIN** and Cheng-Tao CHANG* *Department of Mechanical Engineering, National Cheng Kung University 1 University Road, Tainan 701, Taiwan (R.O.C.) E-mail: [email protected] **Department of Neurology College of Medicine and University Hospital National Cheng Kung University 1 University Road, Tainan 701, Taiwan (R.O.C.) Received: 14 December 2016; Revised: 11 February 2017; Accepted: 6 March 2017 Abstract The biomechanics of peripheral nerve plays an important role in physiology of the healthy and pathology of the diseased such as the diabetic neuropathy, the cauda equine compression and the carpal tunnel syndrome. The other application is recent development of neural prostheses. The goal of this paper is to review researches done by members of Taiwanese Society of Biomechanics on the biomechanics of peripheral nerves in the past two decades. First, a brief introduction of the anatomy of peripheral nerve is given. Then experiment designs and mechanical models for testing the nerves are presented. The material properties ranged from linear elastic to nonlinear visco-elastic and scales ranged from organ level to tissue level are reviewed. Implications of these studies to clinical problems will be addressed and the challenges and future perspective on biomechanics of peripheral nerve are discussed. Key words: Biomechanics, Peripheral nerve, Review, Diabetic mellitus, Cauda equine 1. Introduction Nervous system plays an important role in the human body it can coordinate voluntary and involuntary actions of other tissues and sub-systems. -

Lecture 19: Nerve Synthesis in Vivo (Regeneration)
Nerve synthesis in vivo (regeneration)* 1. Anatomy and function of a peripheral nerve. 2. Experimental parameters for study of induced regeneration. 3. Synthesis of myelinated axons and BM (nerve fibers) 4. Evidence (?) of synthesis of an endoneurium. 5. Synthesis of a nerve trunk (including summary of kinetics of synthesis). 6. Comparative regenerative activity of various reactants. _______ *Tissue and Organ Regeneration in Adults, Yannas, Springer, 2001, Ch. 6. 1. Anatomy and function of a peripheral nerve. I Nervous system = central nervous system (CNS) + peripheral nervous system (PNS) Image: public domain (by Wikipedia User: Persion Poet Gal) Nervous System: CNS and PNS CNS PNS Chamberlain, Yannas, et al., 1998 Landstrom, Aria. “Nerve Regeneration Induced by Collagen-GAG Matrix in Collagen Tubes.” MS Thesis, MIT, 1994. Focus of interest: nerve fibers and axons Nerve fibers comprise axons wrapped in a myelin sheath, itself surrounded by BM (diam. 10-30 μm in rat sciatic nerve). Axons are extensions (long processes) of neurons located in spinal cord. They comprise endoplasmic reticulum and microtubules. 1. Anatomy and function of a peripheral nerve. II Myelinated axons (diam. 1-15 μm) are wrapped in a myelin sheath; nonmyelinated axons also exist. They are the elementary units for conduction of electric signals in the body. Myelin formed by wrapping a Schwann cell membrane many times around axon perimeter. No ECM inside nerve fibers. Myelin sheath is a wrapping of Schwann cell membranes around certain axons. 1. Anatomy and function of a peripheral nerve. III Nonmyelinated axons (diam. <1 μm) function in small pain nerves. Although surrounded by Schwann cells, they lack myelin sheath; Schwann cells are around them but have retained their cytoplasm. -

Kumka's Response to Stecco's Fascial Nomenclature Editorial
Journal of Bodywork & Movement Therapies (2014) 18, 591e598 Available online at www.sciencedirect.com ScienceDirect journal homepage: www.elsevier.com/jbmt FASCIA SCIENCE AND CLINICAL APPLICATIONS: RESPONSE Kumka’s response to Stecco’s fascial nomenclature editorial Myroslava Kumka, MD, PhD* Canadian Memorial Chiropractic College, Department of Anatomy, 6100 Leslie Street, Toronto, ON M2H 3J1, Canada Received 12 May 2014; received in revised form 13 May 2014; accepted 26 June 2014 Why are there so many discussions? response to the direction of various strains and stimuli. (De Zordo et al., 2009) Embedded with a range of mechanore- The clinical importance of fasciae (involvement in patho- ceptors and free nerve endings, it appears fascia has a role in logical conditions, manipulation, treatment) makes the proprioception, muscle tonicity, and pain generation. fascial system a subject of investigation using techniques (Schleip et al., 2005) Pathology and injury of fascia could ranging from direct imaging and dissections to in vitro potentially lead to modification of the entire efficiency of cellular modeling and mathematical algorithms (Chaudhry the locomotor system (van der Wal and Pubmed Exact, 2009). et al., 2008; Langevin et al., 2007). Despite being a topic of growing interest worldwide, This tissue is important for all manual therapists as a controversies still exist regarding the official definition, pain generator and potentially treatable entity through soft terminology, classification and clinical significance of fascia tissue and joint manipulative techniques. (Day et al., 2009) (Langevin et al., 2009; Mirkin, 2008). It is also reportedly treated with therapeutic modalities Lack of consistent terminology has a negative effect on such as therapeutic ultrasound, microcurrent, low level international communication within and outside many laser, acupuncture, and extracorporeal shockwave therapy. -

Nervous System: CNS + PNS
Nervous System: CNS + PNS https://www.webmd.com/brain/ss/slideshow-nervous-system-overview http://paydayloans-mo.com/anatomy-and-physiology-of-central-nervous-system/nervous- system-project-awesome-anatomy-and-physiology-of-central-nervous-system/ A “typical” neuron • Is a cell • Has all the usual cellular components • Usually large nucleus • RER = “Nissl substance/bodies” • Neurites = cell processes Cell body = soma = perikaryon Go to link for animation https://en.wikipedia.org/wiki/Soma_(biology)#/media/File:Neuron_Cell_Body.png Neuron shape categories: found (mainly) in… • Bipolar: organs of special sense • Unipolar: dorsal/posterior root ganglia • Multipolar: everywhere else http://humanphysiology.academy/Neurosciences%202015/Chapter%201/P.1.3p%20Neurone%20Micro.html A simple somatic neural circuit https://www.studyblue.com/notes/hh/cell-bodies-sensory-neurons-gathered-dorsal-root-ganglion/26360419950789715 Neuroglia - “glue” CNS – central nervous system PNS – peripheral nervous system • oligodendrocyte – myelination • Schwann cell – myelination • astrocyte - bind neurons to blood • satellite cell - support nerve vessels; blood brain barrier cell bodies in ganglia • microglia - small, few processes... support & phagocytosis • ependyma - line central cavity: spinal canal & ventricles; ciliated • better circulation of CSF • cell division - growth CNS – central nervous system PNS – peripheral nervous system • nucleus – cluster of cell bodies • ganglion – cluster of cell bodies • tract – bundle of neurites • nerve – bundle of neurites -

The Spinal Cord and Spinal Nerves
14 The Nervous System: The Spinal Cord and Spinal Nerves PowerPoint® Lecture Presentations prepared by Steven Bassett Southeast Community College Lincoln, Nebraska © 2012 Pearson Education, Inc. Introduction • The Central Nervous System (CNS) consists of: • The spinal cord • Integrates and processes information • Can function with the brain • Can function independently of the brain • The brain • Integrates and processes information • Can function with the spinal cord • Can function independently of the spinal cord © 2012 Pearson Education, Inc. Gross Anatomy of the Spinal Cord • Features of the Spinal Cord • 45 cm in length • Passes through the foramen magnum • Extends from the brain to L1 • Consists of: • Cervical region • Thoracic region • Lumbar region • Sacral region • Coccygeal region © 2012 Pearson Education, Inc. Gross Anatomy of the Spinal Cord • Features of the Spinal Cord • Consists of (continued): • Cervical enlargement • Lumbosacral enlargement • Conus medullaris • Cauda equina • Filum terminale: becomes a component of the coccygeal ligament • Posterior and anterior median sulci © 2012 Pearson Education, Inc. Figure 14.1a Gross Anatomy of the Spinal Cord C1 C2 Cervical spinal C3 nerves C4 C5 C 6 Cervical C 7 enlargement C8 T1 T2 T3 T4 T5 T6 T7 Thoracic T8 spinal Posterior nerves T9 median sulcus T10 Lumbosacral T11 enlargement T12 L Conus 1 medullaris L2 Lumbar L3 Inferior spinal tip of nerves spinal cord L4 Cauda equina L5 S1 Sacral spinal S nerves 2 S3 S4 S5 Coccygeal Filum terminale nerve (Co1) (in coccygeal ligament) Superficial anatomy and orientation of the adult spinal cord. The numbers to the left identify the spinal nerves and indicate where the nerve roots leave the vertebral canal. -

685 Mechanisms and Controllers of Eccrine Sweating in Humans Manabu Shiba
[Frontiers in Bioscience S2, 685-696, January 1, 2010] Mechanisms and controllers of eccrine sweating in humans Manabu Shibasaki1, Craig G. Crandall2,3 1Department of Environmental and Life Sciences, Nara Women's University Graduate School of Humanities and Sciences, Nara Japan, 2Department of Internal Medicine, University of Texas Southwestern Medical Center, Dallas, TX, 3Institute for Exercise and Environmental Medicine, Texas Health Presbyterian Hospital of Dallas, TX TABLE OF CONTENTS 1. Abstract 2. Introduction and historical perspective 3. Neural pathway from the brain to sweat gland 4. Non-thermal modulators of sweat rate 4.1. Effect of exercise in modulating sweat rate 4.2. Effect of baroreceptors in modulating sweat rate 4.3. Effect of body fluid status and osmolality on sweat rate 5. Summary 6. Acknowledgement 7. References 1. ABSTRACT 2. INTRODUCTION AND HISTORICAL PERSPECTIVE Human body temperature is regulated within a very narrow range. When exposed to hyperthermic conditions, via Evaporative heat loss is critical for human survival in environmental factors and/or increased metabolism, heat a hot environment, particularly when environmental dissipation becomes vital for survival. In humans, the temperature is higher than skin temperature. Exercise or primary mechanism of heat dissipation, particularly when exposure to a hot environment elevates internal and skin ambient temperature is higher than skin temperature, is temperatures, and subsequently increases sweat rate and evaporative heat loss secondary to sweat secretion from skin blood flow. Historically it was thought that skin eccrine glands. While the primary controller of sweating is temperature was more important than internal temperature the integration between internal and skin temperatures, a in the control of sweating (24, 109).