Heredity Volume 13 Part 3 August 1959
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Trees for Good Fall Color
Selecting Trees for Good Fall Color he fall or spring of the year is an excellent time to plant deciduous trees. Although weather conditions can affect the intensity and actual fall color that will develop on T the leaves from year to year, much of it has to do with the type of tree you plant and its genetic makeup. If you are looking for trees to add to your landscape that produce consistent good fall color from year to year, please refer to the list of some recommended shade and ornamental trees below. This list contains trees that will, in most years, turn a variety of brilliant shades of red, orange, scarlet, and/or purplish-red in autumn. Large trees American Sweetgum Black Gum (Nyssa (Liquidambar styraciflua)- sylvatica)- Yellow to Yellowish-purple-red; orange to scarlet to purple select seedless cultivars. shades. Gingko or Maidenhair Red Maple (Acer rubrum)- (Gingko biloba)- Excellent Colors will vary from red- clear yellow. orange red depending on specific cultivar. Sugar Maple (Acer Katsura Tree (Cercidi- saccharum)- Brilliant phyllum japonicum)- yellow, burnt orange to Yellow-soft apricot-orange. reds. White Oak (Quercus alba)- Common Baldcypress Brown to a rich red to wine (Taxodium distichum)- Soft color. brown to orangish-brown. Linden (Tilia)- Yellow-green American Yellowwood to yellowish. (Cladrastis kentuckea)- Yellow tones. Japanese Zelkova (Zelkova serrata)- Yellow-orange- bronze to deep reddish purples depending on cultivar. Medium Trees American Hornbeam Persian Parrotia or Persian (Carpinus caroliniana)- Ironwood (Parrotia Yellow, orange, red to persica)- Brilliant yellow to reddish-purple: varies orange to scarlet-red. according to cultivar. -

Red, Crimson, Scarlet) a Word Study By: Jonathan Machtemes (Results from KJV Word Search)
Determining Correct Colors in the Early Scriptures (Red, Crimson, Scarlet) a word study by: Jonathan Machtemes (results from KJV word search) Red H119 adm pass appears: 10x translated as: dyed red, red, ruddy related modern words ?: Lat. adamantem "iron" (produces red), many Euro etymologies on "adam/adem" words are in dispute, damage (in the sense of injury, bloodletting) related obry words: H122 adm- red, ruddy H125 admdm- reddish H132 admny- red, ruddy H1818 dm- blood (of living beings, or of grapes) H120 adm- man, "Adam" of gn odn "garden of Eden" H121 adm- "Adam" of gn odn "garden of Eden", city in knon "Canaan" H124 adm- sardius (stone, gem) H123 adum, adm- owu "Esau", due to Gen 25:30. Also related: H130 admy/ adumym- of adum="Edomite", or adumym="Edomites" pl. H127 adme- ground, or perhaps "Adamkind" (Gen 12:3) and so on passed based, in large part, on dm being the root. dm is blood. Blood is a familiar, naturally occurring substance. d, as bizarrely simplistic as it seems to me at times, seems to often draw one's attention specifically to the fact that it appears as a pointed tip. It's used in ahd- one, unity, bd- apart, or dd- nipple, teat, (also dud- beloved, as in "between the teats" or the emotions). a, on the other hand, seems to act most frequently as an augmentative or giver of strength. Perhaps it is a glyph of a bull, or something with horns. Additionally, the d can be seen in words with the meaning or idea of cutting: H1843 do, H1856 dqr, H1417 gdud, or in directional words od, qdm, H1864 drum. -

Lititz Borough Shade Tree List Growth Habit Key Columnar Vase Shaped
Lititz Borough Shade Tree List Growth Habit Key Columnar Vase Shaped Pyramidal Rounded Spreading Small Trees – Mature Height Less Than Thirty Feet (30’) Species Common Name Growth Habit Form Description Crategus Winter King Hawthorn 20-35’ Broad, round head Multi- viridis colored ‘Winter bark, King’ ornamental fruit Prunus x incam Okame Cherry 15-25’ Vase-shaped, Attractive bark; ‘Okame’ becoming rounded with pink flowers in age early spring Syringa reticulata Ivory Silk Tree Lilac 20-25’ Uniform rounded White flowers ‘Ivory Silk’ shape in mid- Summer Medium Trees – Approximate Mature Height of Thirty to Fifty Feet (30-50’) Species Common Name Growth habit Form Description Carpinus American Hornbeam 20-30’ Round spreading, caroliniana native, fall color, to compaction tolerant Gleditsia Thornless Honeylocust 30-40’ Pyramidal Small, lightweight triancanthos var. leaves; Golden yellow inermis fall color; Produces ‘Imperial’ , light shade ‘Skyline’, or ‘Moraine’ Nyssa sylvatica Blackgum 20-30’ Fall foliage includes many shades of yellow, orange, red, purple and scarlet Ostrya American Hophornbeam 25-40’ Pyramidal in youth Attractive bark and virginiana becoming broad hop- like fruit; native to Quercus Sawtooth Oak 35-40’ Pyramidal in youth, Yellow fall color; acutissima becoming rounded attractive bark; to acorns Large Trees – Mature Height Greater Than Fifty Feet (50’) Species Common Name Form Growth Habit Description Acer rubrum Columnar Red Maples 50-60’ Columnar Red flowers, fruit, and ‘Bowhall’ or fall color; native Armstrong Acer rubrum -

Mandevilla Bella.Pdf
New Mandevilla Bella Series Varieties 2020: Bella™ Deep Red This plant makes a big impact at retail and in the garden. Our biggest Bella in the series, this variety is well suited to larger containers and retails best on a trellis. The intense red color makes an impressive display and does not fade as the flowers age. This is a strong vining variety. Bella™ Grande Pink An impressive variety with very large, rosy pink flowers. Bella Grande Pink is well suited to larger containers such as gallons and patio pots. Plants will bloom continuously throughout the summer heat. Bella Grade Pink is colder sensitive and should be kept above 54ºF (12ºC) to avoid leaf drop. Bella™ Grande Red This variety has an improved deep red color, but performs very similar to the original Bella Red. This variety flowers later than others in the series but makes a great season extender item at retail. The growth habit is compact with dense branching, making a solid rounded plant that does not split in production or shipping, ensuring a great retail presentation. Bella™ Hot Pink Well suited for 6-inch or larger containers, Bella Hot Pink is one of the strongest, most dependable varieties in the series. In the first year, Bella Hot Pink has a strong, upright mounded habit perfect for pot production, but will start vining in year two if trellised plants are desired. A European favorite due to its tight well branched structure that does not fall apart as plants gain volume. Bella™ Pink Bella Pink produces a much higher flower count than the other standard pink varieties on the market. -

Program with Speaker Bios (PDF)
A CELEBRATION of the SCARLET & BLACK PROJECT APRIL 29-30, 2021 SCHEDULE OF EVENTS APRIL 29--DAY ONE 11:00 AM Art & Archives of the Scarlet & Black Project 1:00 PM The First Black Alumni: An Exploration of S&B Volume 2 6:00 PM Keynote Address: President Jonathan Holloway APRIL 30--DAY TWO 11:00 AM A Community Project: Scarlet & Black in New Brunswick 1:00 PM The Student Protest Movement at Rutgers: S&B Volume 3 4:00 PM The Legacy of Scarlet & Black: A Conversation with Marisa Fuentes and Deborah Gray White APRIL 29-DAY ONE 11:00 AM ART & ARCHIVES OF THE SCARLET & BLACK PROJECT This panel will explore the images used in the Scarlet & Black Project volumes, the digital archive, and community art projects in New Brunswick. Panelists will discuss how their contributions have helped to shape the images and artwork of the project. Jesse Bayker, Digital Archivist for the Scarlet & Black Project Erika Gorder, University Archivist, Rutgers University Libraries Tracey Johnson, Rutgers University History Department Dan Swern, Producing Director at coLAB Arts APRIL 29-DAY ONE 1:00 PM THE FIRST BLACK ALUMNI: AN EXPLORATION OF SCARLET & BLACK VOLUME 2 This panel will discuss the scholarship of Scarlet & Black, Volume 2: Constructing Race and Gender at Rutgers, 1865-1945. Panelists will discuss how race and gender shaped the experience of some of the first black students to attend Rutgers, such as Paul Robeson and Julia Baxter Bates. Shaun Armste ad, Rutgers University H istory Department Kendra Boyd, Rutgers-Camden University Assistant Professor of History Miya Carey, Binghamton University Presidential Postdoctoral Fellow Bruce Hubbard, Rutgers Class of 1969;Principal of Bruce A. -

Indian Influence on the Chinese Novel the Journey to the West
SINO-PLATONIC PAPERS Number 114 March, 2002 Beyond the Question of the Monkey Imposter: Indian Influence on the Chinese Novel The Journey to the West by Ramnath Subbaraman Victor H. Mair, Editor Sino-Platonic Papers Department of East Asian Languages and Civilizations University of Pennsylvania Philadelphia, PA 19104-6305 USA [email protected] www.sino-platonic.org SINO-PLATONIC PAPERS FOUNDED 1986 Editor-in-Chief VICTOR H. MAIR Associate Editors PAULA ROBERTS MARK SWOFFORD ISSN 2157-9679 (print) 2157-9687 (online) SINO-PLATONIC PAPERS is an occasional series dedicated to making available to specialists and the interested public the results of research that, because of its unconventional or controversial nature, might otherwise go unpublished. The editor-in-chief actively encourages younger, not yet well established, scholars and independent authors to submit manuscripts for consideration. Contributions in any of the major scholarly languages of the world, including romanized modern standard Mandarin (MSM) and Japanese, are acceptable. In special circumstances, papers written in one of the Sinitic topolects (fangyan) may be considered for publication. Although the chief focus of Sino-Platonic Papers is on the intercultural relations of China with other peoples, challenging and creative studies on a wide variety of philological subjects will be entertained. This series is not the place for safe, sober, and stodgy presentations. Sino- Platonic Papers prefers lively work that, while taking reasonable risks to advance the field, capitalizes on brilliant new insights into the development of civilization. Submissions are regularly sent out to be refereed, and extensive editorial suggestions for revision may be offered. Sino-Platonic Papers emphasizes substance over form. -

Color Chart Colorchart
Color Chart AMERICANA ACRYLICS Snow (Titanium) White White Wash Cool White Warm White Light Buttermilk Buttermilk Oyster Beige Antique White Desert Sand Bleached Sand Eggshell Pink Chiffon Baby Blush Cotton Candy Electric Pink Poodleskirt Pink Baby Pink Petal Pink Bubblegum Pink Carousel Pink Royal Fuchsia Wild Berry Peony Pink Boysenberry Pink Dragon Fruit Joyful Pink Razzle Berry Berry Cobbler French Mauve Vintage Pink Terra Coral Blush Pink Coral Scarlet Watermelon Slice Cadmium Red Red Alert Cinnamon Drop True Red Calico Red Cherry Red Tuscan Red Berry Red Santa Red Brilliant Red Primary Red Country Red Tomato Red Naphthol Red Oxblood Burgundy Wine Heritage Brick Alizarin Crimson Deep Burgundy Napa Red Rookwood Red Antique Maroon Mulberry Cranberry Wine Natural Buff Sugared Peach White Peach Warm Beige Coral Cloud Cactus Flower Melon Coral Blush Bright Salmon Peaches 'n Cream Coral Shell Tangerine Bright Orange Jack-O'-Lantern Orange Spiced Pumpkin Tangelo Orange Orange Flame Canyon Orange Warm Sunset Cadmium Orange Dried Clay Persimmon Burnt Orange Georgia Clay Banana Cream Sand Pineapple Sunny Day Lemon Yellow Summer Squash Bright Yellow Cadmium Yellow Yellow Light Golden Yellow Primary Yellow Saffron Yellow Moon Yellow Marigold Golden Straw Yellow Ochre Camel True Ochre Antique Gold Antique Gold Deep Citron Green Margarita Chartreuse Yellow Olive Green Yellow Green Matcha Green Wasabi Green Celery Shoot Antique Green Light Sage Light Lime Pistachio Mint Irish Moss Sweet Mint Sage Mint Mint Julep Green Jadeite Glass Green Tree Jade -

Van Well Nursery
VAN WELL NURSERY Gale Gala® Auvil Early Fuji® Red Jonaprince (Malaga cltv.) (Fuji 218 cltv.) (Red Jonagold) Gale Gala is the “one-pick” Gala strain. A This new Fuji strain matures about six weeks Here’s the red Jonagold that delivers “one- whole tree sport of Royal Gala, this outstand- ahead of standard Red Fuji sports. Hundreds pick” color. The deep red hue shows on all ing strain has all the winning characteristics of acres of Auvil Early Fuji have been planted, the apples, even shaded fruit, at the same of its parent but has deeper red stripes and and it continues to produce a higher percent- time and in many cases the coloring comes a fuller red color. With thousands of trees in age of Washington Extra Fancy grade fruit. earlier than on other strains of Jonagold. Red production, Gale Gala growers report one- Fruit flavor, tree structure, and growth habit Jonaprince has the large size and excellent pick harvests with packouts in the 95% to appear to be identical to other Red Fuji strains. flavor of its parent, Jonagold Prince, plus 100% Washington Extra Fancy grade for color. USPP#10,141. better flavor and storage keeping qualities. USPP#10,114. USPP#11,112. Aztec Fuji® Red Cameo® Adams Apple® Red (DT 2 cltv.) (Dudek cltv.) Delicious (Burchinal cltv.) The new generation Red Fuji is considered A limb sport of Cameo, this Van Well Nursery Adams Apple is just the ticket for fruit growers the reddest blush Fuji sport on the market. offering has all the winning traits of its parent who produce apples in areas where getting This strain is fast becoming the standard by — crisp, juicy, sweet-tart flavor, and outstand- good color is a problem. -

Cultural Practices for Scarlet Royal Jennifer Hashim-Buckey, UCCE Viticulture Farm Advisor and David Ramming, USDA-ARS Research Horticulturist
May-June 2008 Cultural Practices for Scarlet Royal Jennifer Hashim-Buckey, UCCE Viticulture Farm Advisor and David Ramming, USDA-ARS Research Horticulturist Scarlet Royal (US Plant Patent 16,229*) is a mid-season red seedless table grape developed by David Ramming and Ronald Tarailo of the USDA-ARS in Parlier, California. The cultivar, formerly known as B1, was released in 2006 and resulted from the cross of two red seedless USDA selections C33-30 x C51-63. The parentage of Scarlet Royal includes Blackrose, Calmeria, Cardinal, Crimson Seedless, Divizich Early, Italia, Maraville, Muscat of Alexandria, Sultanina and Tafafihi Ahmur. The cultivar produces large (0.8 kg./1.8 lb.), conical-shaped clusters that are medium to well filled. Berries are oval in shape and its appearance is similar to Crimson Seedless. Natural Scarlet Royal berries weigh about 5-7 grams, though berry weight and size is slightly increased when fruit is treated with gibberellic acid. The flesh is firm and meaty and the skin is medium to thick. The flavor is described as sweet and neutral. Scarlet Royal ripens mid-to-late August, filling the harvest window between Flame Seedless and Crimson Seedless. Site Selection and Planting Scarlet Royal is moderately vigorous when planted on its own roots. Rootstock selection should be based on site-specific soil pest or soil chemistry problems. Common rootstock choices for Scarlet Royal have included Freedom and 1103-P, but rootstock effects on fruit yield, quality and vine performance have not yet been evaluated. It is likely that grafting to these and other rootstocks common for table grape production in the San Joaquin Valley will increase vine vigor. -
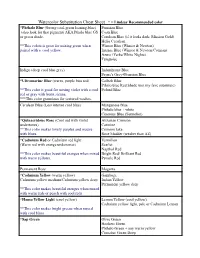
Watercolor Substitution Cheat Sheet * = Lindsay Recommended Color
Watercolor Substitution Cheat Sheet * = Lindsay Recommended color *Phthalo Blue (Strong cool-green leaning-blue) Prussian Blue (also look for that pigment) AKA Pthalo blue GS Cyan Blue or green shade. Cerulean Blue (if it looks dark: Mission Gold) Helio Cerulean **This colors is great for mixing green when Winsor Blue (Winsor & Newton) paired with a cool yellow. Intense Blue (Winsor & Newton/Cotman) Azure (Yarka/White Nights) Turquoise Indigo (deep cool blue grey) Indanthrone Blue Payne's Grey+Prussian Blue *Ultramarine Blue (warm, purple bias red) Colbalt Blue Pthalo blue Red Shade (not my fave substitute) **This color is good for mixing violet with a cool Poland Blue red or gray with burnt sienna. ***This color granulates for textured washes. Cerulean Blue (Less intense cool blue) Manganese Blue Phthalo blue + white Cinerous Blue (Sennelier) *Quinacridone Rose (Cool red with violet Alizarian Crimson undertones) Carmine **This color makes lovely purples and mauve Crimson lake with blues. Rose Madder (weaker than AZ) *Cadmium Red or Cadmium red light Vermilion (Warm red with orange undertones) Scarlet Napthol Red **This color makes beautiful oranges when mixed Bright Red/ Brilliant Red with warm yellows. Pyrrole Red Permanent Rose Magenta *Cadmium Yellow (warm yellow) Gamboge Cadmium yellow medium/Cadmium yellow deep Indian Yellow Permanent yellow deep **This color makes beautiful oranges when mixed with warm reds or peach with cool reds *Hansa Yellow Light (cool yellow) Lemon Yellow (cool yellow) Cadmium yellow light, pale or Cadmium -

Effort to Obtain Data of This Nature in Regard to the Process of Eye of The
THE EYE-PIGMENTARY SYSTEM OF DROSOPHILA VIII. SERIES OF MULTIPLE ALLELES D. J. NOLTE Deportment of Zoology, University of the Witwatersrond Received3.vi.58 THE study of multiple alleles at a locus is of prime importance for research on the theory of the gene—its qualitative and quantitative effects and the relationship between function and mutation. In an effort to obtain data of this nature in regard to the process of eye pigmentation in Drosophila melanogaster nine series of multiple alleles, that is, at the loci of scarlet, vermilion, cinnabar, brown, pink, carna- tion, garnet, raspberry and prune, were investigated for the histology of the four eye-pigment regions and the spectrophotometric properties of the red and brown pigments. These methods have been fully described (Nolte, 1952,1954).These multiple alleles consisted of the following :—scarlct (st) and scarlet-spotted (st'P) ; vermilion (v) and vermilion361 (v361) ; cinnabar (en) and cinnabar3sk (cn3sk) ; brown (bw), brownzb (bw2b), brown-dominant (bwD) and brown4 (bw4); pink (p) and pink-peach (pv) ; carnation (car) and carnationz (carz) garnet (g), garnetz (g2), garnet3 (g3) and garnet4 (g4) ; raspberry (ras), raspberryz (rasz) and raspberry3 (ras3) ; prune (pn) and prune2 (pnz). 2.OBSERVATIONS (I)Phenotype Themacroscopic phenotypes of some of these alleles, in the stocks maintained in this laboratory, differ somewhat from the descriptions as given by Bridges and Brehme (i44), as will duly be noted. The eye-colour of st3 is very like that of the wild-type, with perhaps a pitted effect, i.e., small dark spots, but is not speckled; the combina- tion stsP/st is bright red and not speckled. -

A Study in Scarlet
A Study In Scarlet Arthur Conan Doyle This text is provided to you “as-is” without any warranty. No warranties of any kind, expressed or implied, are made to you as to the text or any medium it may be on, including but not limited to warranties of merchantablity or fitness for a particular purpose. This text was formatted from various free ASCII and HTML variants. See http://sherlock-holm.esfor an electronic form of this text and additional information about it. This text comes from the collection’s version 3.1. Table of contents Part I Mr. Sherlock Holmes . 5 The Science Of Deduction . 8 The Lauriston Garden Mystery . 12 What John Rance Had To Tell. 17 Our Advertisement Brings A Visitor . 20 Tobias Gregson Shows What He Can Do. 23 Light In The Darkness . 27 Part II On The Great Alkali Plain . 35 The Flower Of Utah . 39 John Ferrier Talks With The Prophet . 42 A Flight For Life . 44 The Avenging Angels . 48 A Continuation Of The Reminiscences Of John Watson, M.D. 52 The Conclusion . 57 1 PART I. (Being a reprint from the reminiscences of John H. Watson, M.D., late of the Army Medical Department.) CHAPTER I. Mr.Sherlock Holmes n the year 1878 I took my degree of irresistibly drained. There I stayed for some time at Doctor of Medicine of the University of a private hotel in the Strand, leading a comfortless, London, and proceeded to Netley to go meaningless existence, and spending such money I through the course prescribed for sur- as I had, considerably more freely than I ought.