Effort to Obtain Data of This Nature in Regard to the Process of Eye of The
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
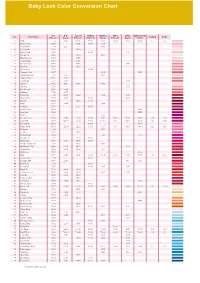
Baby Lock Color Conversion Chart
Tacony_quickguide_37-コピー 05.7.6 9:01 AM ページ 37 Baby Lock Color Conversion Chart R.A. R.A. Isacord Madeira Madeira Sulky Sulky Güetermann No. Color Name Polyester Rayon Polyester Polyneon Rayon Polyester Rayon Dekor Country Embr. 1 Pink 5523 2223 *0180 1921 1121 1224 1108 *4830 155 085 2 Dusty Rose 5675 2155 1816 1108 3 Petal Pink 7701 2255 1015 4 Light Pink 9030 1860 5 Light Coral 9078 1915 1148 6 Ginger Jar 9080 2170 1115 7 Heather Mist 9070 1755 8 Champagne 9063 2051 9 Dark Mauve 9015 2153 1119 10 Heather 9164 2152 11 Neon Pink 5711 1948 12 Comfort Pink 9077 1119 5435 13 Mountain Rose 5795 2373 1315 14 Cherry Pink 5544 2244 15 Carnation 5537 2509 1188 16 Salmon 9073 2553 1840 1018 17 Shrimp 5546 1154 18 Dark Coral 9065 2246 19 Bitteroot 7709 2277 20 Burgundy 5549 2249 2022 1182 1169 21 Warm Wine 5796 2622 1782 1309 22 Russet 5552 2123 1781 23 Plum 9055 2498 1389 24 Maroon 5676 2115 1919 25 Royal Crest 9162 5400 26 Hot Pink 5560 5385 27 Ruby 5797 1183 28 Dark Fuchsia 5804 2504 *2300 1984 1383 1533 1533 *4810 126 107 29 Carmine 5561 *2419 2300 1986 *1081 1511 *1511 5315 158 807 30 Dark Pink 9161 1994 4810 31 Deep Rose 9168 *2508 2520 1721 *1117 1154 1307 *4941 024 086 32 Begonia 5528 1117 33 Azalea 7712 2220 34 Rubine Red 9012 1186 4740 35 Strawberry 5732 2320 1910 36 Devil Red 7706 2507 1906 1986 37 Candy Apple Red 5807 1805 1081 38 Hollyhock Red 9006 2267 1912 1311 39 Toasty Red 9002 2418 1902 1181 40 Wild Fire 5567 4700 41 Red 5678 2505 2101 1637 *1037 1037 *1037 *4740 149 800 42 Jockey Red 5581 1747 43 Radiant Red 5566 2219 561 4731 -

Trees for Good Fall Color
Selecting Trees for Good Fall Color he fall or spring of the year is an excellent time to plant deciduous trees. Although weather conditions can affect the intensity and actual fall color that will develop on T the leaves from year to year, much of it has to do with the type of tree you plant and its genetic makeup. If you are looking for trees to add to your landscape that produce consistent good fall color from year to year, please refer to the list of some recommended shade and ornamental trees below. This list contains trees that will, in most years, turn a variety of brilliant shades of red, orange, scarlet, and/or purplish-red in autumn. Large trees American Sweetgum Black Gum (Nyssa (Liquidambar styraciflua)- sylvatica)- Yellow to Yellowish-purple-red; orange to scarlet to purple select seedless cultivars. shades. Gingko or Maidenhair Red Maple (Acer rubrum)- (Gingko biloba)- Excellent Colors will vary from red- clear yellow. orange red depending on specific cultivar. Sugar Maple (Acer Katsura Tree (Cercidi- saccharum)- Brilliant phyllum japonicum)- yellow, burnt orange to Yellow-soft apricot-orange. reds. White Oak (Quercus alba)- Common Baldcypress Brown to a rich red to wine (Taxodium distichum)- Soft color. brown to orangish-brown. Linden (Tilia)- Yellow-green American Yellowwood to yellowish. (Cladrastis kentuckea)- Yellow tones. Japanese Zelkova (Zelkova serrata)- Yellow-orange- bronze to deep reddish purples depending on cultivar. Medium Trees American Hornbeam Persian Parrotia or Persian (Carpinus caroliniana)- Ironwood (Parrotia Yellow, orange, red to persica)- Brilliant yellow to reddish-purple: varies orange to scarlet-red. according to cultivar. -

Vermilion Category: Alchemy Therasia Von Tux, Barony 1000 Eyes Specific Art Form: Pigment This Is the Sept 2001 Revised Edition (One Small “Oops” Corrected)
Vermilion Category: Alchemy Therasia von Tux, Barony 1000 Eyes specific art form: pigment this is the Sept 2001 revised edition (one small “oops” corrected) Entry contents: 1 vial vermilion/cinnabar (POISON!!! PLEASE DON"T OPEN THE BOTTLE) 1 piece cinnabar 1 copy of Artists' Pigments, for the Gettens et al. article (a copy of Cennini can be found with my "St. George" shield, in the Fine Arts category; copies of Pliny and Theophilus can be found with my silver entry, in the Alchemy category) A piece of cinnabar collected by a friend and on display at the McLaughlin Mine, Napa Co., CA (It's a lot prettier than most of my samples) The first known mention of mercuric sulfide as a mineral is in Theophratus's Lapidary. Theophrastus called it cinnabar, which is the name still in use today. The Romans called it minium, and it was so valuable as a red pigment that the Roman government made it a nationalized monopoly and regulated the price.1 It does need to be said that the Romans used the name minium for any high-quality mineral red, regardless of whether it was cinnabar or hematite.2 Minium was being adultrated with "red lead" as early as Imperial Rome, though it is difficult to determine whether Pliny meant red litharge or lead carbonate. Adulteration continued until the name shifted, and came to mean just red lead by Early Gothic. 3, 4 1 Pliny, Book XXXIII: sections XXXVI-XLI, using the Loeb numbering system. 2 Ibid. 3 Ibid. 4 Though historians would quibble about this, I tend to prefer to the art history divisions of history rather than the academic history divisions. -

Red, Crimson, Scarlet) a Word Study By: Jonathan Machtemes (Results from KJV Word Search)
Determining Correct Colors in the Early Scriptures (Red, Crimson, Scarlet) a word study by: Jonathan Machtemes (results from KJV word search) Red H119 adm pass appears: 10x translated as: dyed red, red, ruddy related modern words ?: Lat. adamantem "iron" (produces red), many Euro etymologies on "adam/adem" words are in dispute, damage (in the sense of injury, bloodletting) related obry words: H122 adm- red, ruddy H125 admdm- reddish H132 admny- red, ruddy H1818 dm- blood (of living beings, or of grapes) H120 adm- man, "Adam" of gn odn "garden of Eden" H121 adm- "Adam" of gn odn "garden of Eden", city in knon "Canaan" H124 adm- sardius (stone, gem) H123 adum, adm- owu "Esau", due to Gen 25:30. Also related: H130 admy/ adumym- of adum="Edomite", or adumym="Edomites" pl. H127 adme- ground, or perhaps "Adamkind" (Gen 12:3) and so on passed based, in large part, on dm being the root. dm is blood. Blood is a familiar, naturally occurring substance. d, as bizarrely simplistic as it seems to me at times, seems to often draw one's attention specifically to the fact that it appears as a pointed tip. It's used in ahd- one, unity, bd- apart, or dd- nipple, teat, (also dud- beloved, as in "between the teats" or the emotions). a, on the other hand, seems to act most frequently as an augmentative or giver of strength. Perhaps it is a glyph of a bull, or something with horns. Additionally, the d can be seen in words with the meaning or idea of cutting: H1843 do, H1856 dqr, H1417 gdud, or in directional words od, qdm, H1864 drum. -

Lititz Borough Shade Tree List Growth Habit Key Columnar Vase Shaped
Lititz Borough Shade Tree List Growth Habit Key Columnar Vase Shaped Pyramidal Rounded Spreading Small Trees – Mature Height Less Than Thirty Feet (30’) Species Common Name Growth Habit Form Description Crategus Winter King Hawthorn 20-35’ Broad, round head Multi- viridis colored ‘Winter bark, King’ ornamental fruit Prunus x incam Okame Cherry 15-25’ Vase-shaped, Attractive bark; ‘Okame’ becoming rounded with pink flowers in age early spring Syringa reticulata Ivory Silk Tree Lilac 20-25’ Uniform rounded White flowers ‘Ivory Silk’ shape in mid- Summer Medium Trees – Approximate Mature Height of Thirty to Fifty Feet (30-50’) Species Common Name Growth habit Form Description Carpinus American Hornbeam 20-30’ Round spreading, caroliniana native, fall color, to compaction tolerant Gleditsia Thornless Honeylocust 30-40’ Pyramidal Small, lightweight triancanthos var. leaves; Golden yellow inermis fall color; Produces ‘Imperial’ , light shade ‘Skyline’, or ‘Moraine’ Nyssa sylvatica Blackgum 20-30’ Fall foliage includes many shades of yellow, orange, red, purple and scarlet Ostrya American Hophornbeam 25-40’ Pyramidal in youth Attractive bark and virginiana becoming broad hop- like fruit; native to Quercus Sawtooth Oak 35-40’ Pyramidal in youth, Yellow fall color; acutissima becoming rounded attractive bark; to acorns Large Trees – Mature Height Greater Than Fifty Feet (50’) Species Common Name Form Growth Habit Description Acer rubrum Columnar Red Maples 50-60’ Columnar Red flowers, fruit, and ‘Bowhall’ or fall color; native Armstrong Acer rubrum -

Mandevilla Bella.Pdf
New Mandevilla Bella Series Varieties 2020: Bella™ Deep Red This plant makes a big impact at retail and in the garden. Our biggest Bella in the series, this variety is well suited to larger containers and retails best on a trellis. The intense red color makes an impressive display and does not fade as the flowers age. This is a strong vining variety. Bella™ Grande Pink An impressive variety with very large, rosy pink flowers. Bella Grande Pink is well suited to larger containers such as gallons and patio pots. Plants will bloom continuously throughout the summer heat. Bella Grade Pink is colder sensitive and should be kept above 54ºF (12ºC) to avoid leaf drop. Bella™ Grande Red This variety has an improved deep red color, but performs very similar to the original Bella Red. This variety flowers later than others in the series but makes a great season extender item at retail. The growth habit is compact with dense branching, making a solid rounded plant that does not split in production or shipping, ensuring a great retail presentation. Bella™ Hot Pink Well suited for 6-inch or larger containers, Bella Hot Pink is one of the strongest, most dependable varieties in the series. In the first year, Bella Hot Pink has a strong, upright mounded habit perfect for pot production, but will start vining in year two if trellised plants are desired. A European favorite due to its tight well branched structure that does not fall apart as plants gain volume. Bella™ Pink Bella Pink produces a much higher flower count than the other standard pink varieties on the market. -

Program with Speaker Bios (PDF)
A CELEBRATION of the SCARLET & BLACK PROJECT APRIL 29-30, 2021 SCHEDULE OF EVENTS APRIL 29--DAY ONE 11:00 AM Art & Archives of the Scarlet & Black Project 1:00 PM The First Black Alumni: An Exploration of S&B Volume 2 6:00 PM Keynote Address: President Jonathan Holloway APRIL 30--DAY TWO 11:00 AM A Community Project: Scarlet & Black in New Brunswick 1:00 PM The Student Protest Movement at Rutgers: S&B Volume 3 4:00 PM The Legacy of Scarlet & Black: A Conversation with Marisa Fuentes and Deborah Gray White APRIL 29-DAY ONE 11:00 AM ART & ARCHIVES OF THE SCARLET & BLACK PROJECT This panel will explore the images used in the Scarlet & Black Project volumes, the digital archive, and community art projects in New Brunswick. Panelists will discuss how their contributions have helped to shape the images and artwork of the project. Jesse Bayker, Digital Archivist for the Scarlet & Black Project Erika Gorder, University Archivist, Rutgers University Libraries Tracey Johnson, Rutgers University History Department Dan Swern, Producing Director at coLAB Arts APRIL 29-DAY ONE 1:00 PM THE FIRST BLACK ALUMNI: AN EXPLORATION OF SCARLET & BLACK VOLUME 2 This panel will discuss the scholarship of Scarlet & Black, Volume 2: Constructing Race and Gender at Rutgers, 1865-1945. Panelists will discuss how race and gender shaped the experience of some of the first black students to attend Rutgers, such as Paul Robeson and Julia Baxter Bates. Shaun Armste ad, Rutgers University H istory Department Kendra Boyd, Rutgers-Camden University Assistant Professor of History Miya Carey, Binghamton University Presidential Postdoctoral Fellow Bruce Hubbard, Rutgers Class of 1969;Principal of Bruce A. -

Indian Influence on the Chinese Novel the Journey to the West
SINO-PLATONIC PAPERS Number 114 March, 2002 Beyond the Question of the Monkey Imposter: Indian Influence on the Chinese Novel The Journey to the West by Ramnath Subbaraman Victor H. Mair, Editor Sino-Platonic Papers Department of East Asian Languages and Civilizations University of Pennsylvania Philadelphia, PA 19104-6305 USA [email protected] www.sino-platonic.org SINO-PLATONIC PAPERS FOUNDED 1986 Editor-in-Chief VICTOR H. MAIR Associate Editors PAULA ROBERTS MARK SWOFFORD ISSN 2157-9679 (print) 2157-9687 (online) SINO-PLATONIC PAPERS is an occasional series dedicated to making available to specialists and the interested public the results of research that, because of its unconventional or controversial nature, might otherwise go unpublished. The editor-in-chief actively encourages younger, not yet well established, scholars and independent authors to submit manuscripts for consideration. Contributions in any of the major scholarly languages of the world, including romanized modern standard Mandarin (MSM) and Japanese, are acceptable. In special circumstances, papers written in one of the Sinitic topolects (fangyan) may be considered for publication. Although the chief focus of Sino-Platonic Papers is on the intercultural relations of China with other peoples, challenging and creative studies on a wide variety of philological subjects will be entertained. This series is not the place for safe, sober, and stodgy presentations. Sino- Platonic Papers prefers lively work that, while taking reasonable risks to advance the field, capitalizes on brilliant new insights into the development of civilization. Submissions are regularly sent out to be refereed, and extensive editorial suggestions for revision may be offered. Sino-Platonic Papers emphasizes substance over form. -

Color Chart Colorchart
Color Chart AMERICANA ACRYLICS Snow (Titanium) White White Wash Cool White Warm White Light Buttermilk Buttermilk Oyster Beige Antique White Desert Sand Bleached Sand Eggshell Pink Chiffon Baby Blush Cotton Candy Electric Pink Poodleskirt Pink Baby Pink Petal Pink Bubblegum Pink Carousel Pink Royal Fuchsia Wild Berry Peony Pink Boysenberry Pink Dragon Fruit Joyful Pink Razzle Berry Berry Cobbler French Mauve Vintage Pink Terra Coral Blush Pink Coral Scarlet Watermelon Slice Cadmium Red Red Alert Cinnamon Drop True Red Calico Red Cherry Red Tuscan Red Berry Red Santa Red Brilliant Red Primary Red Country Red Tomato Red Naphthol Red Oxblood Burgundy Wine Heritage Brick Alizarin Crimson Deep Burgundy Napa Red Rookwood Red Antique Maroon Mulberry Cranberry Wine Natural Buff Sugared Peach White Peach Warm Beige Coral Cloud Cactus Flower Melon Coral Blush Bright Salmon Peaches 'n Cream Coral Shell Tangerine Bright Orange Jack-O'-Lantern Orange Spiced Pumpkin Tangelo Orange Orange Flame Canyon Orange Warm Sunset Cadmium Orange Dried Clay Persimmon Burnt Orange Georgia Clay Banana Cream Sand Pineapple Sunny Day Lemon Yellow Summer Squash Bright Yellow Cadmium Yellow Yellow Light Golden Yellow Primary Yellow Saffron Yellow Moon Yellow Marigold Golden Straw Yellow Ochre Camel True Ochre Antique Gold Antique Gold Deep Citron Green Margarita Chartreuse Yellow Olive Green Yellow Green Matcha Green Wasabi Green Celery Shoot Antique Green Light Sage Light Lime Pistachio Mint Irish Moss Sweet Mint Sage Mint Mint Julep Green Jadeite Glass Green Tree Jade -

A Third Sex-Linked Lethal Factor in Drosophila T
A THIRD SEX-LINKED LETHAL FACTOR IN DROSOPHILA T. H. MORGAN Department of ZoBlogy, Columbia University THREE FIGURES In an experiment (reported in the Jour. Exp. Zool., page 390, November, 1911) in which three sex-linked characters were in- volved, namely, white eyes, vermilion eyes and miniature wings, certain results were obtained involving double crossing-over. The more recent results indicate that double crossing-over was excessive in this experiment.l A white miniature male had been mated to a vermilion pink (‘orange’) female. The daughters were wild type (red, long) and the sons vermilion (long). The two sex- chromosomes of these daughters contain respectively the genes for white (w), red (V), and miniature (m); and for red (W), and vermilion (v), and long (M). All later work has shown the order of the genes here involved is white, vermilion, miniature, as shown in diagram 1. On this basis, the results of the previous experiment in terms of cross-overs would be as follows: Total Per cent Non-cross-overs (vermilion and ver. pink males) 333 74.7 Single cross-overs between white and vermilion (min. and min. pink males). 81 18.2 Single cross-overs between vermilion and min. (verm. min. and ver. min. pink males). 11 2.4 Double cross-overs (wild type and pink males). 21 4.7 Instead of giving fewer double cross-overs than singles, accord- ing to expectation, the number of doubles was twice that of singles, and for numbers as large as these this difference cannot be due to 1 In the analysis on page 392 the large C in ‘sperm no X’ should be small c. -

Genetics and Cytology of <Emphasis Type="Italic">Drosophila Subobscura
GENETICS AND CYTOLOGY OF DROSOPHILA SUBOBSCURA III. TRANSPLANTATION OF EYE-BUDS BETWEEN DROSOPHILA SUBOBSGURA AND DROSOPtlILA MELANOGASTER By J. M. RENDEL, National A~imal B,reeding Research Organization% Institute of Animal Genetics; Edinburgh, AND A. C. E. SULEY, Department of Biomet'ry, University Oollege, London There are in Drosophila subobscura four bl'ight red eye-colour mutants, vermilio~ (chromo- some I, sex-linked), scarlet (chr. II), poppy (chr. IV), and cherry (chr. u An attempt is being made to homologize some of these, and certain other genes, with genes in D. melanogaster, where the phenotypically similar genes, vermilion, cinnabar, cardinal and scarlet do not behave identically in transplantation experiments. As the method depends on the work of Ephrussi & Beadle, the points in their results with D. mclanogaster relevant to this work will be briefly summarized (see Table 1). Implants from larvae of most eye-colour mutants are autonomous with respect to their colour developed in wild-type hosts. But implants from vermilion and cinnabar develop the browner colour of the wild-type host's eye. A genetically vermilion eye also develops the wild-type colour in a cinnabar host, though cinnabar in vermilion is autonomous. Ephrussi & Beadle therefore suggested that two diffusible substances essential to the development of the brown pigment were supplied by the wild-type host. The vermilion mutant lacks one, the v+ substance, which has since been shown to be kymtrenine, and cinnabar lacks the cn+ substance. The behaviour of the reciprocal transplants between the two mutants suggests that the v+ substance is the precursor of the cn + substance. -

Van Well Nursery
VAN WELL NURSERY Gale Gala® Auvil Early Fuji® Red Jonaprince (Malaga cltv.) (Fuji 218 cltv.) (Red Jonagold) Gale Gala is the “one-pick” Gala strain. A This new Fuji strain matures about six weeks Here’s the red Jonagold that delivers “one- whole tree sport of Royal Gala, this outstand- ahead of standard Red Fuji sports. Hundreds pick” color. The deep red hue shows on all ing strain has all the winning characteristics of acres of Auvil Early Fuji have been planted, the apples, even shaded fruit, at the same of its parent but has deeper red stripes and and it continues to produce a higher percent- time and in many cases the coloring comes a fuller red color. With thousands of trees in age of Washington Extra Fancy grade fruit. earlier than on other strains of Jonagold. Red production, Gale Gala growers report one- Fruit flavor, tree structure, and growth habit Jonaprince has the large size and excellent pick harvests with packouts in the 95% to appear to be identical to other Red Fuji strains. flavor of its parent, Jonagold Prince, plus 100% Washington Extra Fancy grade for color. USPP#10,141. better flavor and storage keeping qualities. USPP#10,114. USPP#11,112. Aztec Fuji® Red Cameo® Adams Apple® Red (DT 2 cltv.) (Dudek cltv.) Delicious (Burchinal cltv.) The new generation Red Fuji is considered A limb sport of Cameo, this Van Well Nursery Adams Apple is just the ticket for fruit growers the reddest blush Fuji sport on the market. offering has all the winning traits of its parent who produce apples in areas where getting This strain is fast becoming the standard by — crisp, juicy, sweet-tart flavor, and outstand- good color is a problem.