Medial Temporal Lobe (The Limbic System)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Distinct Representations of Olfactory Information in Different Cortical Centres
LETTER doi:10.1038/nature09868 Distinct representations of olfactory information in different cortical centres Dara L. Sosulski1, Maria Lissitsyna Bloom1{, Tyler Cutforth1{, Richard Axel1 & Sandeep Robert Datta1{ Sensory information is transmitted to the brain where it must be behaviours, but is unlikely to specify innate behaviours. Rather, innate processed to translate stimulus features into appropriate beha- olfactory behaviours are likely to result from the activation of genetically vioural output. In the olfactory system, distributed neural activity determined, stereotyped neural circuits. We have therefore developed a in the nose is converted into a segregated map in the olfactory strategy to trace the projections from identified glomeruli in the olfactory bulb1–3. Here we investigate how this ordered representation is bulb to higher olfactory cortical centres. transformed in higher olfactory centres in mice. We have Mitral and tufted cells that innervate a single glomerulus were developed a tracing strategy to define the neural circuits that labelled by electroporation of tetramethylrhodamine (TMR)-dextran convey information from individual glomeruli in the olfactory under the guidance of a two-photon microscope. This technique labels bulb to the piriform cortex and the cortical amygdala. The spatial mitral and tufted cells that innervate a single glomerulus and is suffi- order in the bulb is discarded in the piriform cortex; axons from ciently robust to allow the identification of axon termini within mul- individual glomeruli project diffusely to the piriform without tiple higher order olfactory centres (Figs 1a–c, 2 and Supplementary apparent spatial preference. In the cortical amygdala, we observe Figs 1–4). Labelling of glomeruli in the olfactory bulbs of mice that broad patches of projections that are spatially stereotyped for indi- express GFP under the control of specific odorant receptor promoters vidual glomeruli. -
Dense Representation of Natural Odorants in the Mouse Olfactory Bulb
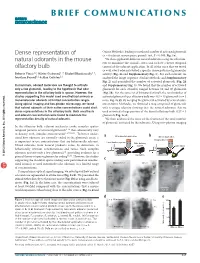
BRIEF COMMUNICATIONS Dense representation of Online Methods), leading to a reduced number of activated glomeruli (n = 6 odorant-mouse pairs, paired t test, P < 0.005; Fig. 1e). natural odorants in the mouse We then applied 40 different natural odorants using the olfactom- eter to minimize the animal’s stress and to have a better temporal olfactory bulb control of the odorant application. In all of the mice that we tested (n = 8), every odorant evoked a specific dense pattern of glomerular Roberto Vincis1,2, Olivier Gschwend1–3, Khaleel Bhaukaurally1–3, activity (Fig. 2a and Supplementary Fig. 1). For each odorant, we Jonathan Beroud1,2 & Alan Carleton1,2 analyzed the image sequence (Online Methods and Supplementary Fig. 2) and quantified the number of activated glomeruli (Fig. 2b In mammals, odorant molecules are thought to activate and Supplementary Fig. 3). We found that the number of activated only a few glomeruli, leading to the hypothesis that odor glomeruli for each stimulus ranged between 10 and 40 glomeruli representation in the olfactory bulb is sparse. However, the (Fig. 2b). For the same set of 30 natural stimuli, the total number of studies supporting this model used anesthetized animals or activated glomeruli per olfactory bulb was 443 ± 15 glomeruli (n = 5 monomolecular odorants at limited concentration ranges. mice; Fig. 2c,d). By merging the glomeruli activated by several odor- Using optical imaging and two-photon microscopy, we found ants (Online Methods), we obtained a map composed of glomeruli that natural odorants at their native concentrations could elicit with a unique identity showing that the natural odorants that we dense representations in the olfactory bulb. -

Functional Organization of Sensory Input to the Olfactory Bulb Glomerulus Analyzed by Two-Photon Calcium Imaging
Functional organization of sensory input to the olfactory bulb glomerulus analyzed by two-photon calcium imaging Matt Wachowiak*, Winfried Denk†, and Rainer W. Friedrich†‡ *Department of Biology, Boston University, 5 Cummington Street, Boston, MA 02215; and †Department of Biomedical Optics, Max Planck Institute for Medical Research, Jahnstrasse 29, D-69120 Heidelberg, Germany Edited by Charles F. Stevens, The Salk Institute for Biological Studies, La Jolla, CA, and approved May 10, 2004 (received for review January 20, 2004) Glomeruli in the olfactory bulb are anatomically discrete modules OSNs distinguished by histochemical markers terminate in dis- receiving input from idiotypic olfactory sensory neurons. To ex- crete subregions of the axonal compartment (36, 37). These data amine the functional organization of sensory inputs to individual raise the possibility that functionally distinct subdivisions exist .glomeruli, we loaded olfactory sensory neurons with a Ca2؉ within a glomerulus indicator and measured odorant-evoked presynaptic Ca2؉ signals Direct measurements of the activity of glomerular afferents within single glomeruli by using two-photon microscopy in anaes- have been difficult because most functional measures of intra- thetized mice. Odorants evoked patterns of discrete Ca2؉ signals glomerular activity lack sufficient spatial resolution or cellular throughout the neuropil of a glomerulus. Across glomeruli, Ca2؉ specificity. Here, we loaded OSNs with a fluorescent Ca2ϩ signals occurred with equal probability in all glomerular regions. indicator and measured patterns of afferent presynaptic activity ؉ Within single glomeruli, the pattern of intraglomerular Ca2 sig- within glomeruli by two-photon Ca2ϩ imaging in anaesthetized nals was indistinguishable for stimuli of different duration, iden- mice. Our data indicate that OSN input to individual glomeruli tity, and concentration. -

Taste and Smell Disorders in Clinical Neurology
TASTE AND SMELL DISORDERS IN CLINICAL NEUROLOGY OUTLINE A. Anatomy and Physiology of the Taste and Smell System B. Quantifying Chemosensory Disturbances C. Common Neurological and Medical Disorders causing Primary Smell Impairment with Secondary Loss of Food Flavors a. Post Traumatic Anosmia b. Medications (prescribed & over the counter) c. Alcohol Abuse d. Neurodegenerative Disorders e. Multiple Sclerosis f. Migraine g. Chronic Medical Disorders (liver and kidney disease, thyroid deficiency, Diabetes). D. Common Neurological and Medical Disorders Causing a Primary Taste disorder with usually Normal Olfactory Function. a. Medications (prescribed and over the counter), b. Toxins (smoking and Radiation Treatments) c. Chronic medical Disorders ( Liver and Kidney Disease, Hypothyroidism, GERD, Diabetes,) d. Neurological Disorders( Bell’s Palsy, Stroke, MS,) e. Intubation during an emergency or for general anesthesia. E. Abnormal Smells and Tastes (Dysosmia and Dysgeusia): Diagnosis and Treatment F. Morbidity of Smell and Taste Impairment. G. Treatment of Smell and Taste Impairment (Education, Counseling ,Changes in Food Preparation) H. Role of Smell Testing in the Diagnosis of Neurodegenerative Disorders 1 BACKGROUND Disorders of taste and smell play a very important role in many neurological conditions such as; head trauma, facial and trigeminal nerve impairment, and many neurodegenerative disorders such as Alzheimer’s, Parkinson Disorders, Lewy Body Disease and Frontal Temporal Dementia. Impaired smell and taste impairs quality of life such as loss of food enjoyment, weight loss or weight gain, decreased appetite and safety concerns such as inability to smell smoke, gas, spoiled food and one’s body odor. Dysosmia and Dysgeusia are very unpleasant disorders that often accompany smell and taste impairments. -

Olfactory Maps, Circuits and Computations
Available online at www.sciencedirect.com ScienceDirect Olfactory maps, circuits and computations Andrew J Giessel and Sandeep Robert Datta Sensory information in the visual, auditory and somatosensory between local positional features to extract information systems is organized topographically, with key sensory features like object identity, depth and motion [4–6]. Unlike the ordered in space across neural sheets. Despite the existence of a small number of continuous sensory parameters that spatially stereotyped map of odor identity within the olfactory characterize vision, audition and touch (such as position, bulb, it is unclear whether the higher olfactory cortex uses frequency and amplitude), olfactory parameter space is topography to organize information about smells. Here, we poorly defined and highly multidimensional [7]. For review recent work on the anatomy, microcircuitry and example, any given monomolecular odorant can be neuromodulation of two higher-order olfactory areas: the described in terms of its functional groups, molecular piriform cortex and the olfactory tubercle. The piriform is an weight, chain length, bond substitution, resonance fre- archicortical region with an extensive local associational network quency or any number of additional chemical descrip- that constructs representations of odor identity. The olfactory tors. Furthermore, olfactory space is inherently tubercle is an extension of the ventral striatum that may use discrete — not only are individual odorants structurally reward-based learning rules to encode odor valence. We argue unique but many of the molecular descriptors typically that in contrast to brain circuits for other sensory modalities, both used for individual odorants (such as functional group or the piriform and the olfactory tubercle largely discard any bond substitution) cannot be mapped continuously in topography present in the bulb and instead use distributive any scheme for chemical space. -

Variants of Olfactory Memory and Their Dependencies on the Hippocampal Formation
The Journal of Neuroscience, February 1995, f5(2): 1162-i 171 Variants of Olfactory Memory and Their Dependencies on the Hippocampal Formation Ursula Sttiubli,’ To-Tam Le,2 and Gary Lynch* ‘Center for Neural Science, New York University, New York, New York 10003 and *Center for the Neurobiology of Learning and Memory, University of California, Irvine, California 92717 Olfactory memory in control rats and in animals with entor- pal pyramidal cells (e.g., Hjorth-Simonsen, 1972; Witter, 1993). hinal cortex lesions was tested in four paradigms: (1) a known Moreover, physiological activity in rat hippocampus becomes correct odor was present in a group of familiar but nonre- synchronized with that in the olfactory bulb and cortex during warded odors, (2) six known correct odors were simulta- odor sampling(Macrides et al., 1982). Theseobservations have neously present in a maze, (3) correct responses required prompted speculationand experimentation concerning the pos- the learning of associations between odors and objects, and sible contributions of the several stagesof the olfactory-hip- (4) six odors, each associated with a choice between two pocampal circuit to the encoding and use of memory. Lesions objects, were presented simultaneously. Control rats had no to the lateral entorhinal cortex, which separatethe hippocampus difficulty with the first problem and avoided repeating se- from its primary source of olfactory input, did not detectably lections in the second; this latter behavior resembles that affect the ability of rats to perform odor discriminations learned reported for spatial mazes but, in the present experiments, prior to surgery although they did disrupt the learning of new was not dependent upon memory for the configuration of discriminations (Staubli et al., 1984, 1986).This result suggested pertinent cues. -

Normal Mentality Associated with a Maldeveloped " Rhinencephalon " by P
J Neurol Neurosurg Psychiatry: first published as 10.1136/jnnp.13.3.191 on 1 August 1950. Downloaded from J. Neurol. Neurosurg. Psychiat., 1950, 13, 191. NORMAL MENTALITY ASSOCIATED WITH A MALDEVELOPED " RHINENCEPHALON " BY P. W. NATHAN and MARION C. SMITH Fronm the Neurological Research Unit of the Medical Research Coulncil, National Hospital. Queen Square, London In 1937 Papez published his views on the Case Report functions of the gyrus fornicatus. He summarized The specimen is the brain of a man, aged 34, who died them as follows. of a chondrosarcoma of the ileum. He was one of a series of cases under investigation in a programme of " It is proposed that the hypothalamus, the anterior thalamic nuclei, the gyrus cinguli, the hippocampus, research concerned with operations for the relief of and their inter-connexions constitute a harmonious pain. As many hours had been devoted to the testing mechanism which may elaborate the functions of of sensation and the discussion of signs and symptoms, central emotion, as well as participate in emotional our knowledge of the patient, which extended over a expression." period of six months, was much greater than that Bilateral temporal lobectomies performed on acquired in most routine cases. Protected by copyright. and Bucy in 1939 gave evidence No abnormality of development had been suspected monkeys by Kiuver during the patient's life, for he came well within normal supporting Papez' general hypothesis. The effects limits from intellectual and psychological points of of extensive lesions of the hippocampus-fornix view. His mother had remained well during pregnancy, system in cats and dogs were reported by Spiegel, and his birth was normal. -

Asymmetric Neurotransmitter Release Enables Rapid Odour Lateralization in Drosophila
SUPPLEMENTARY INFORMATION doi:10.1038/nature11747 SUPPLEMENTARY NOTES Note S1: Light offset produced a precise compensatory turn in the direction opposite to the light-evoked turn (Fig. 1g). As a result, the fly’s net lateral displacement after this compensatory turn was close to zero (Fig. 1i). A train of light pulses produced a larger turn, and the fly’s compensatory turn after light offset was commensurately larger, although less precise (Fig. 1i). This suggests that the fly keeps track of how far it has turned away from its original path, and the accuracy of its estimate depends on how far it has strayed. Note S2: Resting GCaMP fluorescence (F) was not significantly different in the right and left glomeruli (p = 0.67, n = 6, paired t-test). This is expected because each glomerulus contains the axons arising from both the right and left antennae. Although one antenna is removed, the cut axons arising from that antenna are still present in the neuropil. Thus, both right and left glomeruli must contain the same number of GCaMP molecules, and so resting fluorescence is the same. Spontaneous ORN activity evidently makes little contribution to F. What is the mechanistic basis for the significant ipsi/contra difference in F/F? A previous study reported that an ORN plasma membrane marker is two-fold more abundant in the ipsilateral glomerulus as compared to the contralateral glomerulus1. This suggests that the ipsilateral arbor is simply larger than the contralateral arbor of the same ORN. If the properties of ipsi- and contralateral branches are the same, then the ipsi/contra asymmetry in F/F should be roughly proportional to the ipsi/contra asymmetry in arbor size. -

Synaptic Organization of Anterior Olfactory Nucleus Inputs to Piriform Cortex
9414 • The Journal of Neuroscience, December 2, 2020 • 40(49):9414–9425 Systems/Circuits Synaptic Organization of Anterior Olfactory Nucleus Inputs to Piriform Cortex Marco J. Russo,1 Kevin M. Franks,1 Roxanne Oghaz,1 Richard Axel,2 and Steven A. Siegelbaum3 1Department of Neuroscience, Vagelos College of Physicians and Surgeons, Columbia University, New York, New York, 2Mortimer B. Zuckerman Mind Brain Behavior Institute, Department of Neuroscience, Department of Biochemistry and Molecular Biophysics, Howard Hughes Medical Institute, Vagelos College of Physicians & Surgeons, Columbia University, New York, New York, and 3Department of Neuroscience, Kavli Institute for Brain Science, Mortimer B. Zuckerman Mind Brain Behavior Institute, Department of Pharmacology, Vagelos College of Physicians and Surgeons, Columbia University, New York, New York Odors activate distributed ensembles of neurons within the piriform cortex, forming cortical representations of odor thought to be essential to olfactory learning and behaviors. This odor response is driven by direct input from the olfactory bulb, but is also shaped by a dense network of associative or intracortical inputs to piriform, which may enhance or constrain the cort- ical odor representation. With optogenetic techniques, it is possible to functionally isolate defined inputs to piriform cortex and assess their potential to activate or inhibit piriform pyramidal neurons. The anterior olfactory nucleus (AON) receives direct input from the olfactory bulb and sends an associative projection to piriform cortex that has potential roles in the state-dependent processing of olfactory behaviors. Here, we provide a detailed functional assessment of the AON afferents to piriform in male and female C57Bl/6J mice. We confirm that the AON forms glutamatergic excitatory synapses onto piriform pyramidal neurons; and while these inputs are not as strong as piriform recurrent collaterals, they are less constrained by disynaptic inhibition. -

A Cortical Pathway Modulates Sensory Input Into the Olfactory Striatum 3 4 5 Kate A
bioRxiv preprint doi: https://doi.org/10.1101/235291; this version posted December 16, 2017. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. 1 2 A cortical pathway modulates sensory input into the olfactory striatum 3 4 5 Kate A. White1,2,3, Yun-Feng Zhang4, Zhijian Zhang5, Janardhan P. Bhattarai4, Andrew 6 H. Moberly4, Estelle in ‘t Zandt1,2, Huijie Mi6, Xianglian Jia7, Marc V. Fuccillo4, Fuqiang 7 Xu5, Minghong Ma4, Daniel W. Wesson1,2,3* 8 9 1Department of Pharmacology & Therapeutics 10 2Center for Smell and Taste 11 University of Florida 12 1200 Newell Dr.; Gainesville, FL, 32610. U.S.A. 13 3Department of Neurosciences 14 Case Western Reserve University 15 2109 Adelbert Rd.; Cleveland, OH, 44106. U.S.A. 16 4Department of Neuroscience 17 University of Pennsylvania Perelman School of Medicine 18 211 CRB, 415 Curie Blvd; Philadelphia, PA, 19104. U.S.A 19 5Center for Brain Science 20 Wuhan Institute of Physics and Mathematics 21 Chinese Academy of Sciences 22 Wuhan 430071, China 23 6College of Life Sciences 24 Wuhan University 25 Wuhan 430072, China 26 7Shenzhen Institutes of Advanced Technology 27 Chinese Academy of Sciences 28 Shenzhen 518055, China 29 30 *corresponding author; [email protected] 31 RUNNING HEAD: Olfactory striatum input 32 33 Author Contributions: Conceptualization: K.A.W. and D.W.W.; Methodology: K.A.W., Z.Z., F.X., 34 M.M., and D.W.W.; Investigation: K.A.W., Y-F.Z., Z.Z., J.P.B., A.H.M., E.I.Z., H.M., and X.J.; 35 Resources: M.V.F.; Writing – Original Draft: K.A.W., Z.Z., M.M., and D.W.W.; Writing – Review & 36 Editing: all authors; Visualization: K.A.W., Z.Z., Y-F.Z., J.P.B., D.W.W.; Supervision: F.X., M.M., 37 and D.W.W.; Funding Acquisition: K.A.W., F.X., M.M., and D.W.W. -

Olfactory Bulb Glomeruli: External Tufted Cells Intrinsically Burst at Theta Frequency and Are Entrained by Patterned Olfactory Input
1190 • The Journal of Neuroscience, February 4, 2004 • 24(5):1190–1199 Cellular/Molecular Olfactory Bulb Glomeruli: External Tufted Cells Intrinsically Burst at Theta Frequency and Are Entrained by Patterned Olfactory Input Abdallah Hayar, Sergei Karnup, Michael T. Shipley, and Matthew Ennis Department of Anatomy and Neurobiology, Program in Neuroscience, University of Maryland, Baltimore, Maryland 21201 Glomeruli, the initial sites of synaptic processing in the olfactory system, contain at least three types of neurons collectively referred to as juxtaglomerular (JG) neurons. The role of JG neurons in odor processing is poorly understood. We investigated the morphology, spon- taneous, and sensory-evoked activity of one class of JG neurons, external tufted (ET) cells, using whole-cell patch-clamp and extracellular recordings in rat olfactory bulb slices. ET cells have extensive dendrites that ramify within a single glomerulus or, rarely, in two adjacent glomeruli. All ET neurons exhibit spontaneous rhythmic bursts of action potentials (ϳ1–8 bursts/sec). Bursting is intrinsically gener- ated; bursting persisted and became more regular in the presence of ionotropic glutamate and GABA receptor antagonists. Burst fre- quency is voltage dependent; frequency increased at membrane potentials depolarized relative to rest and decreased during membrane potential hyperpolarization. Spontaneous bursting persisted in blockers of calcium channels that eliminated low-threshold calcium spikes (LTS) in ET cells. ET cells have a persistent sodium current available at membrane potentials that generate spontaneous bursting. Internal perfusion with a fast sodium channel blocker eliminated spontaneous bursting but did not block the LTS. These results suggest that persistent sodium channels are essential for spontaneous burst generation in ET cells. -

Tuning and Topography in an Odor Map on the Rat Olfactory Bulb
The Journal of Neuroscience, February 15, 2001, 21(4):1351–1360 Tuning and Topography in an Odor Map on the Rat Olfactory Bulb Markus Meister1,2 and Tobias Bonhoeffer2 1Harvard University, Cambridge, Massachusetts 02138, and 2Max Planck Institute of Neurobiology, D-82152 Munich- Martinsried, Germany The sense of smell originates in a diverse array of receptor centration dependence and yields the effective affinity with neurons, comprising up to 1000 different types. To understand which a glomerulus responds to an odorant. When tested with how these parallel channels encode chemical stimuli, we re- aliphatic molecules of increasing carbon chain length, many corded the responses of glomeruli in the olfactory bulbs of the glomeruli were sharply tuned for one or two adjacent chain anesthetized rat, by optical imaging of intrinsic signals. Odor lengths. Glomeruli with similar tuning properties were located stimulation produced two kinds of optical responses at the near each other, producing a systematic map of molecular surface of the bulb: a broad diffuse component superposed by chain length on the surface of the olfactory bulb. Given local discrete small spots. Histology showed that the spots corre- inhibitory circuits within the olfactory bulb, this can account for spond to individual glomeruli, and that ϳ400 of them can be the observed functional inhibition between related odors. We monitored in this way. Based on its wavelength-dependence, explore several parallels to the function and architecture of the this optical signal appears to derive from changes in light visual system that help interpret the neural representation of scattering during neural activity. Pure odorants generally acti- odors.