Tuning and Topography in an Odor Map on the Rat Olfactory Bulb
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A Rhodopsin Gene Expressed in Photoreceptor Cell R7 of the Drosophila Eye: Homologies with Other Signal-Transducing Molecules
The Journal of Neuroscience, May 1987, 7(5): 1550-I 557 A Rhodopsin Gene Expressed in Photoreceptor Cell R7 of the Drosophila Eye: Homologies with Other Signal-Transducing Molecules Charles S. Zuker, Craig Montell, Kevin Jones, Todd Laverty, and Gerald M. Rubin Department of Biochemistry, University of California, Berkeley, California 94720 We have isolated an opsin gene from D. melanogaster that example, Pak, 1979; Hardie, 1983; Rubin, 1985). The com- is expressed in the ultraviolet-sensitive photoreceptor cell pound eye of Drosophila contains 3 distinct classesof photo- R7 of the Drosophila compound eye. This opsin gene con- receptor cells, Rl-6, R7, and R8, distinguishableby their mor- tains no introns and encodes a 383 amino acid polypeptide phological arrangement and the spectral behavior of their that is approximately 35% homologous to the blue absorbing corresponding visual pigments (reviewed by Hardie, 1983). In ninaE and Rh2 opsins, which are expressed in photoreceptor each of the approximately 800 ommatidia that make up the eye cells RI-6 and R8, respectively. Amino acid homologies be- there are 6 outer (Rl-R6) and 2 central (1 R7 and 1 R8) pho- tween these different opsins and other signal-transducing toreceptor cells (Fig. 1). The photopigments found in the Rl- molecules suggest an important role for the conserved do- R6 cells, the R7 cell, and the R8 cell differ in their absorption mains of rhodopsin in the transduction of extracellular sig- spectra (Harris et al., 1976) most likely becausedifferent opsin nals. genesare expressedin these distinct classesof photoreceptor cells. The 6 peripheral cells (RI-6) contain the major visual Phototransduction, the neuronal excitation processtriggered by pigment, a rhodopsin that absorbsmaximally at 480 nm (Ostroy light, provides an ideal model system for the study of sensory et al., 1974). -

Chemoreception
Senses 5 SENSES live version • discussion • edit lesson • comment • report an error enses are the physiological methods of perception. The senses and their operation, classification, Sand theory are overlapping topics studied by a variety of fields. Sense is a faculty by which outside stimuli are perceived. We experience reality through our senses. A sense is a faculty by which outside stimuli are perceived. Many neurologists disagree about how many senses there actually are due to a broad interpretation of the definition of a sense. Our senses are split into two different groups. Our Exteroceptors detect stimulation from the outsides of our body. For example smell,taste,and equilibrium. The Interoceptors receive stimulation from the inside of our bodies. For instance, blood pressure dropping, changes in the gluclose and Ph levels. Children are generally taught that there are five senses (sight, hearing, touch, smell, taste). However, it is generally agreed that there are at least seven different senses in humans, and a minimum of two more observed in other organisms. Sense can also differ from one person to the next. Take taste for an example, what may taste great to me will taste awful to someone else. This all has to do with how our brains interpret the stimuli that is given. Chemoreception The senses of Gustation (taste) and Olfaction (smell) fall under the category of Chemoreception. Specialized cells act as receptors for certain chemical compounds. As these compounds react with the receptors, an impulse is sent to the brain and is registered as a certain taste or smell. Gustation and Olfaction are chemical senses because the receptors they contain are sensitive to the molecules in the food we eat, along with the air we breath. -

Distinct Representations of Olfactory Information in Different Cortical Centres
LETTER doi:10.1038/nature09868 Distinct representations of olfactory information in different cortical centres Dara L. Sosulski1, Maria Lissitsyna Bloom1{, Tyler Cutforth1{, Richard Axel1 & Sandeep Robert Datta1{ Sensory information is transmitted to the brain where it must be behaviours, but is unlikely to specify innate behaviours. Rather, innate processed to translate stimulus features into appropriate beha- olfactory behaviours are likely to result from the activation of genetically vioural output. In the olfactory system, distributed neural activity determined, stereotyped neural circuits. We have therefore developed a in the nose is converted into a segregated map in the olfactory strategy to trace the projections from identified glomeruli in the olfactory bulb1–3. Here we investigate how this ordered representation is bulb to higher olfactory cortical centres. transformed in higher olfactory centres in mice. We have Mitral and tufted cells that innervate a single glomerulus were developed a tracing strategy to define the neural circuits that labelled by electroporation of tetramethylrhodamine (TMR)-dextran convey information from individual glomeruli in the olfactory under the guidance of a two-photon microscope. This technique labels bulb to the piriform cortex and the cortical amygdala. The spatial mitral and tufted cells that innervate a single glomerulus and is suffi- order in the bulb is discarded in the piriform cortex; axons from ciently robust to allow the identification of axon termini within mul- individual glomeruli project diffusely to the piriform without tiple higher order olfactory centres (Figs 1a–c, 2 and Supplementary apparent spatial preference. In the cortical amygdala, we observe Figs 1–4). Labelling of glomeruli in the olfactory bulbs of mice that broad patches of projections that are spatially stereotyped for indi- express GFP under the control of specific odorant receptor promoters vidual glomeruli. -
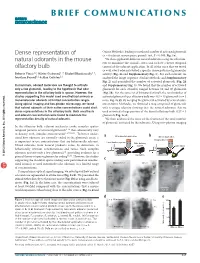
Dense Representation of Natural Odorants in the Mouse Olfactory Bulb
BRIEF COMMUNICATIONS Dense representation of Online Methods), leading to a reduced number of activated glomeruli (n = 6 odorant-mouse pairs, paired t test, P < 0.005; Fig. 1e). natural odorants in the mouse We then applied 40 different natural odorants using the olfactom- eter to minimize the animal’s stress and to have a better temporal olfactory bulb control of the odorant application. In all of the mice that we tested (n = 8), every odorant evoked a specific dense pattern of glomerular Roberto Vincis1,2, Olivier Gschwend1–3, Khaleel Bhaukaurally1–3, activity (Fig. 2a and Supplementary Fig. 1). For each odorant, we Jonathan Beroud1,2 & Alan Carleton1,2 analyzed the image sequence (Online Methods and Supplementary Fig. 2) and quantified the number of activated glomeruli (Fig. 2b In mammals, odorant molecules are thought to activate and Supplementary Fig. 3). We found that the number of activated only a few glomeruli, leading to the hypothesis that odor glomeruli for each stimulus ranged between 10 and 40 glomeruli representation in the olfactory bulb is sparse. However, the (Fig. 2b). For the same set of 30 natural stimuli, the total number of studies supporting this model used anesthetized animals or activated glomeruli per olfactory bulb was 443 ± 15 glomeruli (n = 5 monomolecular odorants at limited concentration ranges. mice; Fig. 2c,d). By merging the glomeruli activated by several odor- Using optical imaging and two-photon microscopy, we found ants (Online Methods), we obtained a map composed of glomeruli that natural odorants at their native concentrations could elicit with a unique identity showing that the natural odorants that we dense representations in the olfactory bulb. -

Functional Organization of Sensory Input to the Olfactory Bulb Glomerulus Analyzed by Two-Photon Calcium Imaging
Functional organization of sensory input to the olfactory bulb glomerulus analyzed by two-photon calcium imaging Matt Wachowiak*, Winfried Denk†, and Rainer W. Friedrich†‡ *Department of Biology, Boston University, 5 Cummington Street, Boston, MA 02215; and †Department of Biomedical Optics, Max Planck Institute for Medical Research, Jahnstrasse 29, D-69120 Heidelberg, Germany Edited by Charles F. Stevens, The Salk Institute for Biological Studies, La Jolla, CA, and approved May 10, 2004 (received for review January 20, 2004) Glomeruli in the olfactory bulb are anatomically discrete modules OSNs distinguished by histochemical markers terminate in dis- receiving input from idiotypic olfactory sensory neurons. To ex- crete subregions of the axonal compartment (36, 37). These data amine the functional organization of sensory inputs to individual raise the possibility that functionally distinct subdivisions exist .glomeruli, we loaded olfactory sensory neurons with a Ca2؉ within a glomerulus indicator and measured odorant-evoked presynaptic Ca2؉ signals Direct measurements of the activity of glomerular afferents within single glomeruli by using two-photon microscopy in anaes- have been difficult because most functional measures of intra- thetized mice. Odorants evoked patterns of discrete Ca2؉ signals glomerular activity lack sufficient spatial resolution or cellular throughout the neuropil of a glomerulus. Across glomeruli, Ca2؉ specificity. Here, we loaded OSNs with a fluorescent Ca2ϩ signals occurred with equal probability in all glomerular regions. indicator and measured patterns of afferent presynaptic activity ؉ Within single glomeruli, the pattern of intraglomerular Ca2 sig- within glomeruli by two-photon Ca2ϩ imaging in anaesthetized nals was indistinguishable for stimuli of different duration, iden- mice. Our data indicate that OSN input to individual glomeruli tity, and concentration. -

Medial Temporal Lobe (The Limbic System)
MEDIAL TEMPORAL LOBE (THE LIMBIC SYSTEM) On the medial surface of the temporal lobe are three structures critical for normal human functioning. From rostral to caudal, they are the olfactory cortex, the amygdala, and the hippocampus. We will look at the anatomy and function of each separately, although they are often grouped together as "the limbic system". A. The olfactory system: The olfactory system actually begins in the roof of the nasal cavity. The olfactory receptors are ciliated epithelial cells with an array of receptors capable of detecting thousands of different odors. However, just as with any sensory system, the receptor neurons themselves do not project to the cerebral hemispheres. Their axons project up through the cribiform plate of the skull to synapse on the dendrites of the mitral cells of the olfactory bulb. The axons of the olfactory receptors make up the elusive cranial nerve I. This fragile tract is susceptible to shearing forces in head trauma, and loss of smell is a surprisingly debilitating injury. Here is an example of a section through olfactory bulb. The olfactory bulb is not a simple relay (something which passively transmits the signal), but is a sophisticated structure in itself. The mitral cell- olfactory neuron synapse is actually within a tangle of axons and dendrites that is called a glomerulus. There is a second cell type tucked around these glomeruli which probably affects how the signal is transmitted. These cells are small and densely packed, which gives them the name "granule cells". However, they bear no relation to the granule cells of the cerebellum or cerebral cortex. -

Intrinsically Different Retinal Progenitor Cells Produce Specific Types Of
PERSPECTIVES These clonal data demonstrated that OPINION RPCs are generally multipotent. However, these data could not determine whether Intrinsically different retinal the variability in clones was due to intrinsic differences among RPCs or extrinsic and/ progenitor cells produce specific or stochastic effects on equivalent RPCs or their progeny. Furthermore, the fates identi- fied within a clone demonstrated an RPC’s types of progeny ‘potential’ but not the ability of an RPC to make a specific cell type at a specific devel- Connie Cepko opmental time or its ‘competence’ (BOX 2). Moreover, although many genes that regu- Abstract | Lineage studies conducted in the retina more than 25 years ago late the development of retinal cell types demonstrated the multipotency of retinal progenitor cells (RPCs). The number have been studied, using the now classical and types of cells produced by individual RPCs, even from a single time point in gain- and loss‑of‑function approaches18,19, development, were found to be highly variable. This raised the question of the precise roles of such regulators in defin- whether this variability was due to intrinsic differences among RPCs or to extrinsic ing an RPC’s competence or potential have not been well elucidated, as most studies and/or stochastic effects on equivalent RPCs or their progeny. Newer lineage have examined the outcome of a perturba- studies that have made use of molecular markers of RPCs, retrovirus-mediated tion on the development of a cell type but lineage analyses of specific RPCs and live imaging have begun to provide answers not the stage and/or cell type in which such a to this question. -

Taste and Smell Disorders in Clinical Neurology
TASTE AND SMELL DISORDERS IN CLINICAL NEUROLOGY OUTLINE A. Anatomy and Physiology of the Taste and Smell System B. Quantifying Chemosensory Disturbances C. Common Neurological and Medical Disorders causing Primary Smell Impairment with Secondary Loss of Food Flavors a. Post Traumatic Anosmia b. Medications (prescribed & over the counter) c. Alcohol Abuse d. Neurodegenerative Disorders e. Multiple Sclerosis f. Migraine g. Chronic Medical Disorders (liver and kidney disease, thyroid deficiency, Diabetes). D. Common Neurological and Medical Disorders Causing a Primary Taste disorder with usually Normal Olfactory Function. a. Medications (prescribed and over the counter), b. Toxins (smoking and Radiation Treatments) c. Chronic medical Disorders ( Liver and Kidney Disease, Hypothyroidism, GERD, Diabetes,) d. Neurological Disorders( Bell’s Palsy, Stroke, MS,) e. Intubation during an emergency or for general anesthesia. E. Abnormal Smells and Tastes (Dysosmia and Dysgeusia): Diagnosis and Treatment F. Morbidity of Smell and Taste Impairment. G. Treatment of Smell and Taste Impairment (Education, Counseling ,Changes in Food Preparation) H. Role of Smell Testing in the Diagnosis of Neurodegenerative Disorders 1 BACKGROUND Disorders of taste and smell play a very important role in many neurological conditions such as; head trauma, facial and trigeminal nerve impairment, and many neurodegenerative disorders such as Alzheimer’s, Parkinson Disorders, Lewy Body Disease and Frontal Temporal Dementia. Impaired smell and taste impairs quality of life such as loss of food enjoyment, weight loss or weight gain, decreased appetite and safety concerns such as inability to smell smoke, gas, spoiled food and one’s body odor. Dysosmia and Dysgeusia are very unpleasant disorders that often accompany smell and taste impairments. -

“Análisis De Los Receptores Tirosina Quinasa ALK, RET Y ROS En Los Adenocarcinomas Nasosinusales”
Universidad de Oviedo Programa de Doctorado “Biomedicina y Oncología Molecular” “Análisis de los receptores tirosina quinasa ALK, RET y ROS en los adenocarcinomas nasosinusales” TESIS DOCTORAL Esteban Reinaldo Pacheco Coronel 20/02/2017 Universidad de Oviedo Programa de Doctorado “Biomedicina y Oncología Molecular” TESIS DOCTORAL “Análisis de los receptores tirosina quinasa ALK, RET y ROS en los adenocarcinomas nasosinusales” Autor: Directores: Esteban Reinaldo José Luís Llorente Pendás Pacheco Coronel Mario Hermsen Dedicatoria A mi familia y amigos, por estar siempre a mi lado y apoyarme en cada momento. Agradecimientos A José Luis por brindarme la oportunidad de trabajar en un tema ambicioso y muy interesante, por los buenos consejos y el tiempo invertido para que este proyecto salga adelante. A Mario, por su valiosa colaboración en el laboratorio, en el análisis de muestras e interpretación de resultados, sus enseñanzas de las diferentes técnicas aplicadas y manejo en el laboratorio; sus consejos sobre la metodología, resultados y conclusiones del proyecto. A mis compañeros del servicio de Otorrinolaringología del Hospital Central de Asturias por sus enseñanzas y el trabajo en equipo. A los compañeros del Instituto Universitario de Oncología del Principado de Asturias. Por que gracias a su trabajo hemos aprendido y desarrollado técnicas importantes para la elaboración de este proyecto. 1 ANTECEDENTES ............................................................................... 1 1.1 Introducción ................................................................................... -

Notch-Signaling in Retinal Regeneration and Müller Glial Plasticity
Notch-Signaling in Retinal Regeneration and Müller glial Plasticity DISSERTATION Presented in Partial Fulfillment of the Requirements for the Degree Doctor of Philosophy in the Graduate School of The Ohio State University By Kanika Ghai, MS Neuroscience Graduate Studies Program The Ohio State University 2009 Dissertation Committee: Dr. Andy J Fischer, Advisor Dr. Heithem El-Hodiri Dr. Susan Cole Dr. Paul Henion Copyright by Kanika Ghai 2009 ABSTRACT Eye diseases such as blindness, age-related macular degeneration (AMD), diabetic retinopathy and glaucoma are highly prevalent in the developed world, especially in a rapidly aging population. These sight-threatening diseases all involve the progressive loss of cells from the retina, the light-sensing neural tissue that lines the back of the eye. Thus, developing strategies to replace dying retinal cells or prolonging neuronal survival is essential to preserving sight. In this regard, cell-based therapies hold great potential as a treatment for retinal diseases. One strategy is to stimulate cells within the retina to produce new neurons. This dissertation elucidates the properties of the primary support cell in the chicken retina, known as the Müller glia, which have recently been shown to possess stem-cell like properties, with the potential to form new neurons in damaged retinas. However, the mechanisms that govern this stem-cell like ability are less well understood. In order to better understand these properties, we analyze the role of one of the key developmental processes, i.e., the Notch-Signaling Pathway in regulating proliferative, neuroprotective and regenerative properties of Müller glia and bestow them with this plasticity. -

Asymmetric Neurotransmitter Release Enables Rapid Odour Lateralization in Drosophila
SUPPLEMENTARY INFORMATION doi:10.1038/nature11747 SUPPLEMENTARY NOTES Note S1: Light offset produced a precise compensatory turn in the direction opposite to the light-evoked turn (Fig. 1g). As a result, the fly’s net lateral displacement after this compensatory turn was close to zero (Fig. 1i). A train of light pulses produced a larger turn, and the fly’s compensatory turn after light offset was commensurately larger, although less precise (Fig. 1i). This suggests that the fly keeps track of how far it has turned away from its original path, and the accuracy of its estimate depends on how far it has strayed. Note S2: Resting GCaMP fluorescence (F) was not significantly different in the right and left glomeruli (p = 0.67, n = 6, paired t-test). This is expected because each glomerulus contains the axons arising from both the right and left antennae. Although one antenna is removed, the cut axons arising from that antenna are still present in the neuropil. Thus, both right and left glomeruli must contain the same number of GCaMP molecules, and so resting fluorescence is the same. Spontaneous ORN activity evidently makes little contribution to F. What is the mechanistic basis for the significant ipsi/contra difference in F/F? A previous study reported that an ORN plasma membrane marker is two-fold more abundant in the ipsilateral glomerulus as compared to the contralateral glomerulus1. This suggests that the ipsilateral arbor is simply larger than the contralateral arbor of the same ORN. If the properties of ipsi- and contralateral branches are the same, then the ipsi/contra asymmetry in F/F should be roughly proportional to the ipsi/contra asymmetry in arbor size. -

Anatomy and Physiology of the Afferent Visual System
Handbook of Clinical Neurology, Vol. 102 (3rd series) Neuro-ophthalmology C. Kennard and R.J. Leigh, Editors # 2011 Elsevier B.V. All rights reserved Chapter 1 Anatomy and physiology of the afferent visual system SASHANK PRASAD 1* AND STEVEN L. GALETTA 2 1Division of Neuro-ophthalmology, Department of Neurology, Brigham and Womens Hospital, Harvard Medical School, Boston, MA, USA 2Neuro-ophthalmology Division, Department of Neurology, Hospital of the University of Pennsylvania, Philadelphia, PA, USA INTRODUCTION light without distortion (Maurice, 1970). The tear–air interface and cornea contribute more to the focusing Visual processing poses an enormous computational of light than the lens does; unlike the lens, however, the challenge for the brain, which has evolved highly focusing power of the cornea is fixed. The ciliary mus- organized and efficient neural systems to meet these cles dynamically adjust the shape of the lens in order demands. In primates, approximately 55% of the cortex to focus light optimally from varying distances upon is specialized for visual processing (compared to 3% for the retina (accommodation). The total amount of light auditory processing and 11% for somatosensory pro- reaching the retina is controlled by regulation of the cessing) (Felleman and Van Essen, 1991). Over the past pupil aperture. Ultimately, the visual image becomes several decades there has been an explosion in scientific projected upside-down and backwards on to the retina understanding of these complex pathways and net- (Fishman, 1973). works. Detailed knowledge of the anatomy of the visual The majority of the blood supply to structures of the system, in combination with skilled examination, allows eye arrives via the ophthalmic artery, which is the first precise localization of neuropathological processes.