Dieciousness in Thalictrum Dasycarpum.*
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

FLORA from FĂRĂGĂU AREA (MUREŞ COUNTY) AS POTENTIAL SOURCE of MEDICINAL PLANTS Silvia OROIAN1*, Mihaela SĂMĂRGHIŢAN2
ISSN: 2601 – 6141, ISSN-L: 2601 – 6141 Acta Biologica Marisiensis 2018, 1(1): 60-70 ORIGINAL PAPER FLORA FROM FĂRĂGĂU AREA (MUREŞ COUNTY) AS POTENTIAL SOURCE OF MEDICINAL PLANTS Silvia OROIAN1*, Mihaela SĂMĂRGHIŢAN2 1Department of Pharmaceutical Botany, University of Medicine and Pharmacy of Tîrgu Mureş, Romania 2Mureş County Museum, Department of Natural Sciences, Tîrgu Mureş, Romania *Correspondence: Silvia OROIAN [email protected] Received: 2 July 2018; Accepted: 9 July 2018; Published: 15 July 2018 Abstract The aim of this study was to identify a potential source of medicinal plant from Transylvanian Plain. Also, the paper provides information about the hayfields floral richness, a great scientific value for Romania and Europe. The study of the flora was carried out in several stages: 2005-2008, 2013, 2017-2018. In the studied area, 397 taxa were identified, distributed in 82 families with therapeutic potential, represented by 164 medical taxa, 37 of them being in the European Pharmacopoeia 8.5. The study reveals that most plants contain: volatile oils (13.41%), tannins (12.19%), flavonoids (9.75%), mucilages (8.53%) etc. This plants can be used in the treatment of various human disorders: disorders of the digestive system, respiratory system, skin disorders, muscular and skeletal systems, genitourinary system, in gynaecological disorders, cardiovascular, and central nervous sistem disorders. In the study plants protected by law at European and national level were identified: Echium maculatum, Cephalaria radiata, Crambe tataria, Narcissus poeticus ssp. radiiflorus, Salvia nutans, Iris aphylla, Orchis morio, Orchis tridentata, Adonis vernalis, Dictamnus albus, Hammarbya paludosa etc. Keywords: Fărăgău, medicinal plants, human disease, Mureş County 1. -

Purple Meadow Rue, Thalictrum Dasycarpum
Purple Meadow Rue, Thalictrum dasycarpum Purple Meadow Rue is a delightful native perennial, 2-6 ft. high, that features blue- green lacy foliage (somewhat resembling columbine) and wiry-branched sprays of tiny, white flowers which bloom for 2-3 weeks in early summer (late May-July). The mass effect of the tiny flowers is quite showy! Emerging in loose, large clumps of soft, almost feathery sprays, the flowerheads can be a foot or more long, but have no true petals. Mostly dioecious (male/female flowers on separate plants), the male flowers are more fluffy with numerous yellow, thread-like stamens that droop out and become entangled. The female flowers are pollinated by wind and develop into spindle-shaped seeds that turn brown at maturity. A member of the buttercup family (Ranunculaceae), Purple Meadow Rue has an upright central stem that is unbranched below, but becomes branched above. The stems are usually purplish. The leaves are hairy beneath, alternate and compound: divided into numerous, three-lobed leaflets which become smaller as they ascend the stems. In the wild, Purple Meadow Rue is found in deciduous riparian woods, damp thickets and ravines, and moist prairies. Bees are sometimes attracted to the abundant pollen of the male flowers. However, this plant is not cross-pollinated by insects as the female flowers do not have nectar. There are a select few insect species that feed on this plant, including some aphids and caterpillars of certain moths. White-Tailed Deer browse the foliage sparingly. This species is currently being researched, because its seeds are one of the best sources of thalicarpine, a chemical with cancer-controlling properties. -

Appendix 1. Species Summaries and Element Occurrence Records For
Appendix 1. Species summaries and element occurrence records for Campanula uniflora, Cymopterus evertii, Descurainia torulosa Helictotrichon hookeri, Papaver kluanense, and Thalictrum alpinum in the Carter Mountain Area of Critical Environmental Concern UYC»4ING NATURAL DIVERSITY DATABASE - The Nature Conservancy Key to Selected Fields in the Element Occurrence Database SNAME - The scientific name, in Latin, used in Wyoming (may be different in another state). SCOMNAME - The common name used in Wyoming. GRANK - The global rank assigned by TNC's network of Heritage Programs, based on world-wide distribution and threats. Ranks vary from G1, very rare or greatly threatened, through GS, common and secure. SRANK- The state rank assigned by each state Heritage Program, based on distribution within the state. Again, these ranks vary from S1, very rare or threatened, through SS, common and secure. These ranks may be different from state to state depending on the range of the taxon in each state. WYPLANT L!Sl-- WYNDD maintains a state lis~ where plants of special concern in the state appear on List 1, High Priority (rare, threatened, or endangered), List 2, Medium Priority, or List 3, Low Priority. PRECISION - The degree of refinement for an occurrence when it is mapped on a USGS quad; S=within seconds, M=within minutes, G=general (somewhere on the cited quad). COUNTYNAME - The county where the occurrence is located. QUADNAME - The name of the USGS 7.5 minute quad. MARG NUM indicates the dot on the map at the WYNDD office. LAT, LONG - Central latitude and longitude of the location if known from mapping procedures. -

Seed Plant Phylogeny: Demise of the Anthophyte Hypothesis? Michael J
bb10c06.qxd 02/29/2000 04:18 Page R106 R106 Dispatch Seed plant phylogeny: Demise of the anthophyte hypothesis? Michael J. Donoghue* and James A. Doyle† Recent molecular phylogenetic studies indicate, The first suggestions that Gnetales are related to surprisingly, that Gnetales are related to conifers, or angiosperms were based on several obvious morphological even derived from them, and that no other extant seed similarities — vessels in the wood, net-veined leaves in plants are closely related to angiosperms. Are these Gnetum, and reproductive organs made up of simple, results believable? Is this a clash between molecules unisexual, flower-like structures, which some considered and morphology? evolutionary precursors of the flowers of wind-pollinated Amentiferae, but others viewed as being reduced from Addresses: *Harvard University Herbaria, 22 Divinity Avenue, Cambridge, Massachusetts 02138, USA. †Section of Evolution and more complex flowers in the common ancestor of Ecology, University of California, Davis, California 95616, USA. angiosperms, Gnetales and Mesozoic Bennettitales [1]. E-mail: [email protected] These ideas went into eclipse with evidence that simple [email protected] flowers really are a derived, rather than primitive, feature Current Biology 2000, 10:R106–R109 of the Amentiferae, and that vessels arose independently in angiosperms and Gnetales. Vessels in angiosperms 0960-9822/00/$ – see front matter seem derived from tracheids with scalariform pits, whereas © 2000 Elsevier Science Ltd. All rights reserved. in Gnetales they resemble tracheids with circular bor- dered pits, as in conifers. Gnetales are also like conifers in These are exciting times for those interested in plant lacking scalariform pitting in the primary xylem, and in evolution. -

Common Name: TRAILING MEADOWRUE Scientific Name: Thalictrum Debile Buckley Other Commonly Used Names: Southern Meadow-Rue Previ
Common Name: TRAILING MEADOWRUE Scientific Name: Thalictrum debile Buckley Other Commonly Used Names: southern meadow-rue Previously Used Scientific Names: Thalictrum arkansanum Boivin, Thalictrum texanum (Gray) Small Family: Ranunculaceae (buttercup) Rarity Ranks: G2/S1 State Legal Status: Threatened Federal Legal Status: none Federal Wetland Status: FAC Description: Perennial herb with slender, sprawling stems 4 - 18 inches (10 - 45 cm) long. Leaves alternate, each leaf divided into many rounded, 3- or 5-lobed leaflets up to ½ inch (0.5 - 1.5 cm) long and wide, upper surface green, lower surface pale. Female and male flowers are on separate plants, in open, branching clusters on slender stalks up to half the length of the plant. Flowers lack petals but have 4 pale green or purple sepals, about inch (1 - 3.5 mm) long; female flowers produce 3 - 5 tiny, ribbed fruits; male flowers contain 12 - 18 yellow stamens. Similar Species: Early meadowrue (Thalictrum dioicum) also has female and male flowers on separate plants. It is 12 - 32 inches (30 - 80 cm) tall with erect stems; its largest leaflets are wider than ½ inch (1.3 cm). Related Rare Species: Leatherleaf meadowrue (Thalictrum coriaceum, Special Concern) occurs in open, deciduous woods in Union County. It has erect stems up to 5 feet (150 cm) tall, and its leaves are divided into many leaflets with pointed lobes or teeth. It flowers May–July. Also see Cooley’s meadowrue (T. cooleyi) on this web site. Habitat: Forested floodplains over limestone. Life History: Trailing meadowrue is a perennial herb with a tuberous rootstock; it dies back to the rootstock each winter and resprouts in the spring, producing one or more stems per tuber cluster. -
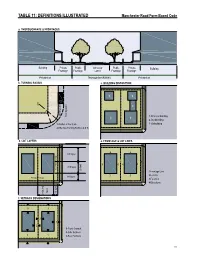
Manchester Road Redevelopment District: Form-Based Code
TaBle 11: deFiniTionS illuSTraTed manchester road Form-Based Code a. ThoroughFare & FronTageS Building Private Public Vehicular Public Private Building Frontage Frontage Lanes Frontage Frontage Private lot Thoroughfare (r.o.w.) Private lot b. Turning radiuS c. Building diSPoSiTion 3 3 2 2 1 Parking Lane Moving Lane 1- Principal Building 1 1 2- Backbuilding 1-Radius at the Curb 3- Outbuilding 2-Effective Turning Radius (± 8 ft) d. loT LAYERS e. FronTage & loT lineS 4 3rd layer 4 2 1 4 4 4 3 2nd layer Secondary Frontage 20 feet 1-Frontage Line 2-Lot Line 1st layer 3 3 Principal Frontage 3-Facades 1 1 4-Elevations layer 1st layer 2nd & 3rd & 2nd f. SeTBaCk deSignaTionS 3 3 2 1 2 1-Front Setback 2-Side Setback 1 1 3-Rear Setback 111 Manchester Road Form-Based Code ARTICLE 9. APPENDIX MATERIALS MBG Kemper Center PlantFinder About PlantFinder List of Gardens Visit Gardens Alphabetical List Common Names Search E-Mail Questions Menu Quick Links Home Page Your Plant Search Results Kemper Blog PlantFinder Please Note: The following plants all meet your search criteria. This list is not necessarily a list of recommended plants to grow, however. Please read about each PF Search Manchesterplant. Some may Road be invasive Form-Based in your area or may Code have undesirable characteristics such as above averageTab insect LEor disease 11: problems. NATIVE PLANT LIST Pests Plants of Merit Missouri Native Plant List provided by the Missouri Botanical Garden PlantFinder http://www.mobot.org/gardeninghelp/plantfinder Master Search Search limited to: Missouri Natives Search Tips Scientific Name Scientific Name Common NameCommon Name Height (ft.) ZoneZone GardeningHelp (ft.) Acer negundo box elder 30-50 2-10 Acer rubrum red maple 40-70 3-9 Acer saccharinum silver maple 50-80 3-9 Titles Acer saccharum sugar maple 40-80 3-8 Acer saccharum subsp. -

Anthropogenic Fire, Vegetation Structure and Ethnobotanical Uses in an Alpine Shrubland of Nepal’S Himalaya
International Journal of Wildland Fire 2020, 29, 201–214 © IAWF 2020 doi: 10.1071/WF19098_AC p.1 Supplementary material Anthropogenic fire, vegetation structure and ethnobotanical uses in an alpine shrubland of Nepal’s Himalaya Asha PaudelA,B,F, Scott H. MarkwithB, Katie KoncharC, Mani ShresthaD,E and Suresh K. GhimireA,F ACentral Department of Botany, Tribhuvan University, Kathmandu, 44618, Nepal. BDepartment of Geosciences, Florida Atlantic University, 777 Glades Road, Boca Raton, FL, 33431, USA. C1334 Jackson Street, Tallahassee, FL, 32301, USA. DSchool of Media and Communication, RMIT University, Melbourne, Vic. 3001, Australia. EFaculty of Information Technology, Monash University, Melbourne, Vic. 3800, Australia. FCorresponding authors. Email: [email protected], [email protected] International Journal of Wildland Fire © IAWF 2020 doi: 10.1071/WF19098_AC p.2 Table S1: List of species recorded from burned and unburned habitat patches in Lauribina Hill, Langtang National Park, Nepal, their elevation range, palatability, ethnobotanical use and chorotype. Scientific name and family Habitat1 Elevation Palatability Major ethnobotanical use4 Chorotype5 range2 for herbivores3 Agrostis pilosula Trin. (Poaceae) BR+UB 2000-4600 P - IN Aletris pauciflora (Klotzsch) Hand.-Mazz. BR+UB 2500-4900 D Medicine: aerial parts in lung and liver disorders [2,3]. PH (Nartheciaceae) Anaphalis nepalensis var. monocephala (DC.) BR+UB 4100-5500 N Medicine: aerial parts in fever, indigestion, inner bleeding PH Hand.-Mazz. (Asteraceae) and swellings [4]. Other: dried plant used as fire catcher [1]. Anemone demissa Hook.f. & Thomson UB 2700-5000 N Medicine: whole plant applied to blisters and wart [2]. PH (Ranunculaceae) Anemone rupestris Wall. ex Hook.f. & Thomson BR+UB 3500-5000 P - PH (Ranunculaceae) Anemone smithiana Lauener & Panigrahi BR+UB 3000-4500 - - PH (Ranunculaceae) Arenaria bryophylla Fernald (Caryophyllaceae) BR 4200-5900 N Medicine: whole plant to control inflammation/pain of PH kidney and burning sensation of bladder/urine tract [5]. -

Ecological Sorting of Vascular Plant Classes During the Paleozoic Evolutionary Radiation
i1 Ecological Sorting of Vascular Plant Classes During the Paleozoic Evolutionary Radiation William A. DiMichele, William E. Stein, and Richard M. Bateman DiMichele, W.A., Stein, W.E., and Bateman, R.M. 2001. Ecological sorting of vascular plant classes during the Paleozoic evolutionary radiation. In: W.D. Allmon and D.J. Bottjer, eds. Evolutionary Paleoecology: The Ecological Context of Macroevolutionary Change. Columbia University Press, New York. pp. 285-335 THE DISTINCTIVE BODY PLANS of vascular plants (lycopsids, ferns, sphenopsids, seed plants), corresponding roughly to traditional Linnean classes, originated in a radiation that began in the late Middle Devonian and ended in the Early Carboniferous. This relatively brief radiation followed a long period in the Silurian and Early Devonian during wrhich morphological complexity accrued slowly and preceded evolutionary diversifications con- fined within major body-plan themes during the Carboniferous. During the Middle Devonian-Early Carboniferous morphological radiation, the major class-level clades also became differentiated ecologically: Lycopsids were cen- tered in wetlands, seed plants in terra firma environments, sphenopsids in aggradational habitats, and ferns in disturbed environments. The strong con- gruence of phylogenetic pattern, morphological differentiation, and clade- level ecological distributions characterizes plant ecological and evolutionary dynamics throughout much of the late Paleozoic. In this study, we explore the phylogenetic relationships and realized ecomorphospace of reconstructed whole plants (or composite whole plants), representing each of the major body-plan clades, and examine the degree of overlap of these patterns with each other and with patterns of environmental distribution. We conclude that 285 286 EVOLUTIONARY PALEOECOLOGY ecological incumbency was a major factor circumscribing and channeling the course of early diversification events: events that profoundly affected the structure and composition of modern plant communities. -

Ranunculaceae – Buttercup Family
RANUNCULACEAE – BUTTERCUP FAMILY Plant: mostly herbs, some woody vines or shrubs Stem: Root: Leaves: mostly alternate, sometimes opposite or whorled or basal; lobed or not lobed; if lobed then most often palmately, but occasionally pinnately, sometimes finely dissected – highly variable, sometimes even on the same plant; with or without stipules Flowers: mostly perfect, some dioecious; sepals 3-6, commonly 5; petals vary in number (3-23) but often 5, petals may be lacking and sepals are showy; stamens few to many; ovary superior, carpels few to very many, pistils one to many Fruit: mostly a dry capsule, seeds small, may be oily; rarely a berry Other: large family, sometimes confused with members of the Rose family (5 petals); Dicotyledons Group Genera: 60+ genera; locally Actaea (baneberry), Anemone (anemone or windflower), Aquilegia (columbine), Clematis, Isopyrum, Hepatica, Hydrastis, Ranunuculus (buttercup or crowfoot), Thalictrum (meadow-rue) WARNING – family descriptions are only a layman’s guide and should not be used as definitive Flower Morphology in the This is a large family often based on 5’s but Ranunculaceae (Buttercup Family) exceptions occur Examples of common genera White Baneberry [Doll’s-Eyes] Yellow Marsh Marigold [Cowslip] Goldenseal [Yellowroot] Actaea pachypoda Ell. Carolina [Wild Blue] Larkspur Caltha palustris L. var. palustris Delphinium carolinianum Walter Hydrastis canadensis L. Swamp Leather Flower [Eastern] False Rue Anemone Clematis crispa L. Devil-In-The-Bush [Love American Wood Anemone Enemion biternatum Raf. -In-A-Mist] Anemone quinquefolia L. [Isopyrum biternatum] Nigella damascena L. (Introduced) Doubtful [Rocket; Garden] Knight's-Spur [Larkspur] Round-lobed Hepatica [Liverleaf] Tall Buttercup Hepatica nobilis Schreber var. -

Ancient Noeggerathialean Reveals the Seed Plant Sister Group Diversified Alongside the Primary Seed Plant Radiation
Ancient noeggerathialean reveals the seed plant sister group diversified alongside the primary seed plant radiation Jun Wanga,b,c,1, Jason Hiltond,e, Hermann W. Pfefferkornf, Shijun Wangg, Yi Zhangh, Jiri Beki, Josef Pšenickaˇ j, Leyla J. Seyfullahk, and David Dilcherl,m,1 aState Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing 210008, China; bCenter for Excellence in Life and Paleoenvironment, Chinese Academy of Sciences, Nanjing 210008, China; cUniversity of Chinese Academy of Sciences, Shijingshan District, Beijing 100049, China; dSchool of Geography, Earth and Environmental Sciences, University of Birmingham, Edgbaston, Birmingham B15 2TT, United Kingdom; eBirmingham Institute of Forest Research, University of Birmingham, Edgbaston, Birmingham B15 2TT, United Kingdom; fDepartment of Earth and Environmental Science, University of Pennsylvania, Philadelphia, PA 19104-6316; gState Key Laboratory of Systematic and Evolutionary Botany, Institute of Botany, Chinese Academy of Sciences, Xiangshan, Beijing 100093, China; hCollege of Paleontology, Shenyang Normal University, Key Laboratory for Evolution of Past Life in Northeast Asia, Ministry of Natural Resources, Shenyang 110034, China; iDepartment of Palaeobiology and Palaeoecology, Institute of Geology v.v.i., Academy of Sciences of the Czech Republic, 165 00 Praha 6, Czech Republic; jCentre of Palaeobiodiversity, West Bohemian Museum in Plzen, 301 36 Plzen, Czech Republic; kDepartment of Paleontology, Geozentrum, University of Vienna, 1090 Vienna, Austria; lIndiana Geological and Water Survey, Bloomington, IN 47404; and mDepartment of Geology and Atmospheric Science, Indiana University, Bloomington, IN 47405 Contributed by David Dilcher, September 10, 2020 (sent for review July 2, 2020; reviewed by Melanie Devore and Gregory J. -
Native Plants North Georgia
Native Plants of North Georgia A photo guide for plant enthusiasts Mickey P. Cummings · The University of Georgia® · College of Agricultural and Environmental Sciences · Cooperative Extension CONTENTS Plants in this guide are arranged by bloom time, and are listed alphabetically within each bloom period. Introduction ................................................................................3 Blood Root .........................................................................5 Common Cinquefoil ...........................................................5 Robin’s-Plantain ..................................................................6 Spring Beauty .....................................................................6 Star Chickweed ..................................................................7 Toothwort ..........................................................................7 Early AprilEarly Trout Lily .............................................................................8 Blue Cohosh .......................................................................9 Carolina Silverbell ...............................................................9 Common Blue Violet .........................................................10 Doll’s Eye, White Baneberry ...............................................10 Dutchman’s Breeches ........................................................11 Dwarf Crested Iris .............................................................11 False Solomon’s Seal .........................................................12 -

Dispersal Effects on Species Distribution and Diversity Across Multiple Scales in the Southern Appalachian Mixed Mesophytic Flora
DISPERSAL EFFECTS ON SPECIES DISTRIBUTION AND DIVERSITY ACROSS MULTIPLE SCALES IN THE SOUTHERN APPALACHIAN MIXED MESOPHYTIC FLORA Samantha M. Tessel A dissertation submitted to the faculty at the University of North Carolina at Chapel Hill in partial fulfillment of the requirements for the degree of Doctor of Philosophy in Ecology in the Curriculum for the Environment and Ecology. Chapel Hill 2017 Approved by: Peter S. White Robert K. Peet Alan S. Weakley Allen H. Hurlbert Dean L. Urban ©2017 Samantha M. Tessel ALL RIGHTS RESERVED ii ABSTRACT Samantha M. Tessel: Dispersal effects on species distribution and diversity across multiple scales in the southern Appalachian mixed mesophytic flora (Under the direction of Peter S. White) Seed and spore dispersal play important roles in the spatial distribution of plant species and communities. Though dispersal processes are often thought to be more important at larger spatial scales, the distribution patterns of species and plant communities even at small scales can be determined, at least in part, by dispersal. I studied the influence of dispersal in southern Appalachian mixed mesophytic forests by categorizing species by dispersal morphology and by using spatial pattern and habitat connectivity as predictors of species distribution and community composition. All vascular plant species were recorded at three nested sample scales (10000, 1000, and 100 m2), on plots with varying levels of habitat connectivity across the Great Smoky Mountains National Park. Models predicting species distributions generally had higher predictive power when incorporating spatial pattern and connectivity, particularly at small scales. Despite wide variation in performance, models of locally dispersing species (species without adaptations to dispersal by wind or vertebrates) were most frequently improved by the addition of spatial predictors.