Lquat Arctic Alpine Plants
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Iconic Bees: 12 Reports on UK Bee Species
Iconic Bees: 12 reports on UK bee species Bees are vital to the ecology of the UK and provide significant social and economic benefits through crop pollination and maintaining the character of the landscape. Recent years have seen substantial declines in many species of bees within the UK. This report takes a closer look at how 12 ‘iconic’ bee species are faring in each English region, as well as Wales, Northern Ireland and Scotland. Authors Rebecca L. Evans and Simon G. Potts, University of Reading. Photo: © Amelia Collins Contents 1 Summary 2 East England Sea-aster Mining Bee 6 East Midlands Large Garden Bumblebee 10 London Buff-tailed Bumblebee 14 North East Bilberry Bumblebee 18 North West Wall Mason Bee 22 Northern Ireland Northern Colletes 26 Scotland Great Yellow Bumblebee 30 South East England Potter Flower Bee 34 South West England Scabious Bee 38 Wales Large Mason Bee 42 West Midlands Long-horned Bee 46 Yorkshire Tormentil Mining Bee Through collating information on the 12 iconic bee species, common themes have Summary emerged on the causes of decline, and the actions that can be taken to help reverse it. The most pervasive causes of bee species decline are to be found in the way our countryside has changed in the past 60 years. Intensification of grazing regimes, an increase in pesticide use, loss of biodiverse field margins and hedgerows, the trend towards sterile monoculture, insensitive development and the sprawl of towns and cities are the main factors in this. I agree with the need for a comprehensive Bee Action Plan led by the UK Government in order to counteract these causes of decline, as called for by Friends of the Earth. -

Plateau Mountain Plant List
Plateau ALBERTA WILDERNESS Mountain – Plant list ASSOCIATION Plants of Plateau Mountain Ecological Reserve (Plateau Mountain Ecological Reserve Management Plan – Alberta Environment) Alpine Anemone Anemone drummondii Drummond's Rock Cress Alpine Arnica Arnica angustifolia Drummond's Rush Alpine Bistort Polygonum viviparum Dwarf Birch Betula glandulosa Alpine Blue Grass Poa alpina Dwarf Bitter root Alpine Everlasting Antennaria sp. Dwarf Hawk's Beard Alpine Fleabane Erigeron pallens Dwarf Sawwort Saussurea nuda Alpine Forget-me not Myosotis alpestris Dwarf saw-wort Saussurea nuda Alpine Goldenrod Solidago multiradianta Dwarf Scouring-rush Equisetum scirpoides Alpine Hawkweed Early Blue Grass Alpine Milk-vetch Astragalus alpinus Early Blue Violet Viola adunca Alpine mouse-eared chickweed Cerastium beeringianum Early Cinquefoil Potertilla concinna Alpine Speedwell Veronica alpina Elephant's head Pedicularis groenlandica Alpine speedwell Veronica alpina Elephant's-head Lousewort Pedicualaris groenlandica Alpine Timothy Elgelmann Spruce Picea engelmanii American Vetch Entire-leaved Groundsel Androsace Androsace chamaejasme Everlasting Antennaria luzuloides Arctic Aster False Dandelion Agoseris aurantiaca Arctic Blue Glass Felwort Gentianela amarella Arctic Butterweed Few-flowered Milk-vetch Astragalus sp. Balsam Groundsel Few-seeded Whitlow-grass Draba oligosperma Balsam Poplar Populus balsamifera Field Chickweed Stellaria sp. Barratt's Willow Fireweed Locoweed Oxytropis sp. Bearberry Arctostaphylos uva-ursi Flame-colored Lousewort Pedicularis -

Potentilla Spp.)-The Five Finger Weeds 1
r Intriguing World of Weeds iiiiiiiiiaiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiii Cinquefoils (Potentilla spp.)-The Five Finger Weeds 1 LARRY W. MITICH2 INTRODUCTION In 1753 Linneaus named the genus Potentilla in his Species Plantarum (4). The common name five finger is m cd frequently for this group of plants ( 18, 29). The genus, in the rose family (Rosaceae), is composed of about 500 north temperate species (50 in North America, 75 Euro pean species) of mostly boreal herbs and shrubs. Indeed, Potentilla extends far into arctic regions (22, 29). How ever, a few species are south temperate. And although less common, some species are also found in alpine and high 11,ountain regions of the tropics and South America; P. anserinoides Lehm. is a New Zealand native (27). Cinquefoil, which means five leaves, is an old herb, full of mystery and magic, which matches the charm Rough cinquefoil, Potentilla norvegica L. of its name. The plant protects its frag ile blooms in bad weather by contract ing the leaves so that they curve over the liver in humans (29). It was prescribed as a tea or in and shelter the flower (11). Cinquefoil wine for diarrhea, leukorrhea, kidney stones, arthritis, was credited with supernatural powers, cramps, and reducing fever (22). However, in recent times and was an essential ingredient in love divination. Accord the roots are being used for a gargle and mouthwash (11). ing to Alice Elizabeth Bacon, frogs liked to sit on this In America the outer root bark of creeping cinquefoil (P. plant-"the toad will be much under Sage, frogs will be in reptans L.) is used to stop nosebleeds. -

Field Release of the Gall Mite, Aceria Drabae
United States Department of Field release of the gall mite, Agriculture Aceria drabae (Acari: Marketing and Regulatory Eriophyidae), for classical Programs biological control of hoary Animal and Plant Health Inspection cress (Lepidium draba L., Service Lepidium chalapense L., and Lepidium appelianum Al- Shehbaz) (Brassicaceae), in the contiguous United States. Environmental Assessment, January 2018 Field release of the gall mite, Aceria drabae (Acari: Eriophyidae), for classical biological control of hoary cress (Lepidium draba L., Lepidium chalapense L., and Lepidium appelianum Al-Shehbaz) (Brassicaceae), in the contiguous United States. Environmental Assessment, January 2018 Agency Contact: Colin D. Stewart, Assistant Director Pests, Pathogens, and Biocontrol Permits Plant Protection and Quarantine Animal and Plant Health Inspection Service U.S. Department of Agriculture 4700 River Rd., Unit 133 Riverdale, MD 20737 Non-Discrimination Policy The U.S. Department of Agriculture (USDA) prohibits discrimination against its customers, employees, and applicants for employment on the bases of race, color, national origin, age, disability, sex, gender identity, religion, reprisal, and where applicable, political beliefs, marital status, familial or parental status, sexual orientation, or all or part of an individual's income is derived from any public assistance program, or protected genetic information in employment or in any program or activity conducted or funded by the Department. (Not all prohibited bases will apply to all programs and/or employment activities.) To File an Employment Complaint If you wish to file an employment complaint, you must contact your agency's EEO Counselor (PDF) within 45 days of the date of the alleged discriminatory act, event, or in the case of a personnel action. -

Maynooth University Publications 2018 Peer Reviewed Journal Name Department Affiliations Journal Pub Title Authors
Maynooth University Publications 2018 Peer Reviewed Journal Name Department Affiliations Journal Pub Title Authors DR LAURA ANDERSON MUSIC - MUSIC AND LETTERS Sonic ‘detheatricalisation’: Jean Cocteau, Film Music, and 'Les Parents terribles' Laura Anderson DR ALBERTO ARRIBAS SOCIOLOGY - Current Sociology Reframing the public sociology debate: Towards collaborative and decolonial praxis. Arribas Lozano, A. Knowledge co-production with social movement networks. Redefining grassroots DR ALBERTO ARRIBAS SOCIOLOGY - Social Movement Studies politics, rethinking research. Arribas Lozano, A. Migraciones, acción colectiva y colonialidad del saber en el campo académico DR ALBERTO ARRIBAS SOCIOLOGY - - español: los y las migrantes como sujetos políticos invisibles/invisibilizados. Arribas Lozano, A. PROFESSOR LAUREN ARRINGTON ENGLISH - Yeats Annual Fighting Spirits: Yeats, Pound and The Winding Stair (1929) Arrington, Lauren PROFESSOR LAUREN ARRINGTON ENGLISH - Irish Political Studies The Blindness of Hindsight: Irish and British Poets Reflect on Early Fascist Italy Arrington, Lauren Histoire et Civilisation du Livre. Revue « Rebelle malgré lui » - récits de réconciliation et de réintégration dans les MS MONIKA BARGET ARTS & HUMANITIES INSTITUTE - Internationale biographies politiques britanniques du XVIIIe siècle Barget, Monika DR OLIVER BARTLETT LAW - European Journal of Risk Regulation Reforming the Regulation on Spirit Drinks – an Example of Better Regulation? O Bartlett DR OLIVER BARTLETT LAW - European Law Journal Power, Policy Ideas and Paternalism in Non-Communicable Disease Prevention O Bartlett DR BERNHARD BAUER SCHOOL OF CELTIC STUDIES - Zeitschrift für celtische Philologie The Story of the Monk and the Devil Bernhard Bauer The Aspergillus nidulans pyruvate dehydrogenase kinases are essential to integrate Ries L.;de Assis L.;Rodrigues F.;Caldana C.;Rocha M.;Malavazi I.;Bayram DR OZGUR BAYRAM BIOLOGY - G3 (Bethesda, Md.) carbon source metabolism Ö.;Goldman G. -

Maine's Endangered and Threatened Plants
University of Southern Maine USM Digital Commons Maine Collection 1990 Maine's Endangered and Threatened Plants Maine State Planning Office Follow this and additional works at: https://digitalcommons.usm.maine.edu/me_collection Part of the Biodiversity Commons, Botany Commons, Ecology and Evolutionary Biology Commons, Forest Biology Commons, Forest Management Commons, Other Forestry and Forest Sciences Commons, Plant Biology Commons, and the Weed Science Commons Recommended Citation Maine State Planning Office, "Maine's Endangered and Threatened Plants" (1990). Maine Collection. 49. https://digitalcommons.usm.maine.edu/me_collection/49 This Book is brought to you for free and open access by USM Digital Commons. It has been accepted for inclusion in Maine Collection by an authorized administrator of USM Digital Commons. For more information, please contact [email protected]. BACKGROUND and PURPOSE In an effort to encourage the protection of native Maine plants that are naturally reduced or low in number, the State Planning Office has compiled a list of endangered and threatened plants. Of Maine's approximately 1500 native vascular plant species, 155, or about 10%, are included on the Official List of Maine's Plants that are Endangered or Threatened. Of the species on the list, three are also listed at the federal level. The U.S. Fish and Wildlife Service. has des·ignated the Furbish's Lousewort (Pedicularis furbishiae) and Small Whorled Pogonia (lsotria medeoloides) as Endangered species and the Prairie White-fringed Orchid (Platanthera leucophaea) as Threatened. Listing rare plants of a particular state or region is a process rather than an isolated and finite event. -
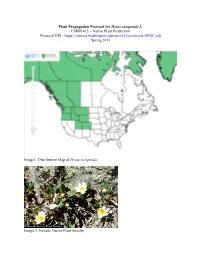
Plant Propagation Protocol for Dryas Octopetala L. ESRM 412 – Native Plant Production Protocol URL
Plant Propagation Protocol for Dryas octopetala L. ESRM 412 – Native Plant Production Protocol URL: https://courses.washington.edu/esrm412/protocols/DROC.pdf Spring 2015 Image1. Distribution Map of Dryas octopetala Image 2. Nevada Native Plant Society TAXONOMY Plant Family [3] Scientific Name Rosaceae Common Name Rose Family Species Scientific Name Scientific Name Dryas octopetala L. [3] Varieties * Dryas octopetala var. angustifolia C.L. Hitchc. [3] Dryas octopetala var. hookeriana (Juz) Hulten Sub-species Dryas octopetala f. argentea (Blytt) Hulten * Dryas octopetala subsp. alaskensis (Porsild) Hulten [3] Cultivar Common Synonym(s) Dradetum octopetalae Keiner Common Name(s) White mountain-avens, Eightpetal mountain-avens, Mountain dryas Species Code (as per USDA Plants DROC database) GENERAL INFORMATION Geographical range Alaska, Washington, Oregon, Colorado Alpine regions in the Pacific Northwest N. Cascades, and Rocky Mountain ranges Ecological distribution Mid-montane to Alpine zone Climate and elevation range Elevation: 3,500 m [6] 100 m and less Climate: sites with low snow cover on calcareous or basic soils [6] Local habitat and abundance Full sun Dry, well-drained, sandy or gravelly soils Spreads rapidly [6] Dominant or co-dominant species within its range [6] Associated species: Dwarf willow Plant strategy type / successional Nitrogen fixer: forms association with Frankia [5] stage Colonizer of barren slopes at high elevations [5] Plant characteristics Forb/herb, Shrub, Subshrub Perrennial Forms mats up to 3 ft. wide and 8 in. tall. [1] 1 cream or white flower at the end of each 2-8 inch leafless flower stalk. [2] Flowers bloom June-July Summer fruits fluffy and feathery Seeds are wind-dispersed [7] Leaves are oval-shaped, leathery with rounded teeth and a white underside. -

The Flora of Jan Mayen
NORSK POLARINSTITUTT SKRIFTER NR. 130 JOHANNES LID THE FLORA OF JAN MAYEN IlJustrated by DAGNY TANDE LID or1(f t ett} NORSK POLARINSTITUTT OSLO 1964 DET KONGELIGE DEPARTEMENT FOR INDUSTRI OG HÅNDVERK NORSK POLARINSTITUTT Observatoriegt. l, Oslo, Norway Short account of the publications of Norsk Polarinstitutt The two series, Norsk Polarinstitutt - SKRIFTER and Norsk Polarinstitutt - MEDDELELSER, were taken over from the institution Norges Svalbard- og Ishavs undersøkelser (NSIU), which was incorporated in Norsk Polarinstitutt when this was founded in 1948. A third series, Norsk Polarinstitutt - ÅRBOK, is published with one volum(� per year. SKRIFTER includes scientific papers, published in English, French or German. MEDDELELSER comprises shortcr papers, often being reprillts from other publi cations. They generally have a more popular form and are mostly published in Norwegian. SKRIFTER has previously been published under various tides: Nos. 1-11. Resultater av De norske statsunderstuttede Spitsbergen-ekspe. ditioner. No 12. Skrifter om Svalbard og Nordishavet. Nos. 13-81. Skrifter om Svalbard og Ishavet. 82-89. Norges Svalbard- og Ishavs-undersøkelser. Skrifter. 90- • Norsk Polarinstitutt Skrifter. In addition a special series is published: NORWEGIAN-BRITISH-SWEDISH ANTARCTIC EXPEDITION, 1949-52. SCIENTIFIC RESULTS. This series will comprise six volumes, four of which are now completed. Hydrographic and topographic surveys make an important part of the work carried out by Norsk Polarinstitutt. A list of the published charts and maps is printed on p. 3 and 4 of this cover. A complete list of publications, charts and maps is obtainable on request. ÅRBØKER Årbok 1960. 1962. Kr.lS.00. Årbok 1961. 1962. Kr. 24.00. -

Buchbesprechungen 247-296 ©Verein Zur Erforschung Der Flora Österreichs; Download Unter
ZOBODAT - www.zobodat.at Zoologisch-Botanische Datenbank/Zoological-Botanical Database Digitale Literatur/Digital Literature Zeitschrift/Journal: Neilreichia - Zeitschrift für Pflanzensystematik und Floristik Österreichs Jahr/Year: 2006 Band/Volume: 4 Autor(en)/Author(s): Mrkvicka Alexander Ch., Fischer Manfred Adalbert, Schneeweiß Gerald M., Raabe Uwe Artikel/Article: Buchbesprechungen 247-296 ©Verein zur Erforschung der Flora Österreichs; download unter www.biologiezentrum.at Neilreichia 4: 247–297 (2006) Buchbesprechungen Arndt KÄSTNER, Eckehart J. JÄGER & Rudolf SCHUBERT, 2001: Handbuch der Se- getalpflanzen Mitteleuropas. Unter Mitarbeit von Uwe BRAUN, Günter FEYERABEND, Gerhard KARRER, Doris SEIDEL, Franz TIETZE, Klaus WERNER. – Wien & New York: Springer. – X + 609 pp.; 32 × 25 cm; fest gebunden. – ISBN 3-211-83562-8. – Preis: 177, – €. Dieses imposante Kompendium – wohl das umfangreichste Werk zu diesem Thema – behandelt praktisch alle Aspekte der reinen und angewandten Botanik rund um die Ackerbeikräuter. Es entstand in der Hauptsache aufgrund jahrzehntelanger Forschungs- arbeiten am Institut für Geobotanik der Universität Halle über Ökologie und Verbrei- tung der Segetalpflanzen. Im Zentrum des Werkes stehen 182 Arten, die ausführlich behandelt werden, wobei deren eindrucksvolle und umfassende „Porträt-Zeichnungen“ und genaue Verbreitungskarten am wichtigsten sind. Der „Allgemeine“ Teil („I.“) beginnt mit der Erläuterung einiger (vor allem morpholo- gischer, ökologischer, chorologischer und zoologischer) Fachausdrücke, darauf -

Dryas Octopetala Subsp. Alaskensis: Potential for a Hardy, Novel Flower
Dryas octopetala subsp. alaskensis: Potential for a Hardy, Novel Flower Eamon Spanier-Scanlon, Student of Plant Science University of Minnesota, St. Paul, MN 5/13/2019 Executive Summary Dryas octopetala, Mountain Avens, is a new sprawling variety of ground cover which may become available in northern climates near you. This extremely cold-tolerant plant will spread easily in your garden or within a larger pot with other plants. With petite white flowers blooming late in the spring, and placed gingerly upon thin red stalks, their beauty persists throughout the summer. This perennial ground cover will return year after year, regardless of the harshest winters. Purchase a few and watch as they creep around your yard, or simply buy one and propagate your own, with ease, through cuttings. The northern consumer may often feel left out by the lack of perennial flowering plants, but with this new species you can soon become the envy of your friends residing in the south. Minimal input is required for these beautiful white flowers to flourish in your domain. Pick up your own in a local floral retailer and fall in love over and over, each spring, as they bound back to life. I. Introduction A. Species The distribution of the genus Dryas is worldwide; they are often found in harsh areas such as arctic and tundra regions (Springer et al., 2019). Dryas octopetala is currently on the market as a decorative, flowering plant, although subsp. alaskensis is not. It can be purchased as a seed from multiple vendors, coming in such names as “Mountain Evans,” available through GeoSeed company (https://www.geoseed.com). -

Astereae, Asteraceae) Using Molecular Phylogeny of ITS
Turkish Journal of Botany Turk J Bot (2015) 39: 808-824 http://journals.tubitak.gov.tr/botany/ © TÜBİTAK Research Article doi:10.3906/bot-1410-12 Relationships and generic delimitation of Eurasian genera of the subtribe Asterinae (Astereae, Asteraceae) using molecular phylogeny of ITS 1, 2,3 4 Elena KOROLYUK *, Alexey MAKUNIN , Tatiana MATVEEVA 1 Central Siberian Botanical Garden, Siberian Branch of Russian Academy of Sciences, Novosibirsk, Russia 2 Institute of Molecular and Cell Biology, Siberian Branch of Russian Academy of Sciences, Novosibirsk, Russia 3 Theodosius Dobzhansky Center for Genome Bioinformatics, Saint Petersburg State University, Saint Petersburg, Russia 4 Department of Genetics & Biotechnology, Saint Petersburg State University, Saint Petersburg, Russia Received: 12.10.2014 Accepted/Published Online: 02.04.2015 Printed: 30.09.2015 Abstract: The subtribe Asterinae (Astereae, Asteraceae) includes highly variable, often polyploid species. Recent findings based on molecular methods led to revision of its volume. However, most of these studies lacked species from Eurasia, where a lot of previous taxonomic treatments of the subtribe exist. In this study we used molecular phylogenetics methods with internal transcribed spacer (ITS) as a marker to resolve evolutionary relations between representatives of the subtribe Asterinae from Siberia, Kazakhstan, and the European part of Russia. Our reconstruction revealed that a clade including all Asterinae species is paraphyletic. Inside this clade, there are species with unresolved basal positions, for example Erigeron flaccidus and its relatives. Moreover, several well-supported groups exist: group of the genera Galatella, Crinitaria, Linosyris, and Tripolium; group of species of North American origin; and three related groups of Eurasian species: typical Eurasian asters, Heteropappus group (genera Heteropappus, Kalimeris), and Asterothamnus group (genera Asterothamnus, Rhinactinidia). -

Arctic National Wildlife Refuge Volume 2
Appendix F Species List Appendix F: Species List F. Species List F.1 Lists The following list and three tables denote the bird, mammal, fish, and plant species known to occur in Arctic National Wildlife Refuge (Arctic Refuge, Refuge). F.1.1 Birds of Arctic Refuge A total of 201 bird species have been recorded on Arctic Refuge. This list describes their status and abundance. Many birds migrate outside of the Refuge in the winter, so unless otherwise noted, the information is for spring, summer, or fall. Bird names and taxonomic classification follow American Ornithologists' Union (1998). F.1.1.1 Definitions of classifications used Regions of the Refuge . Coastal Plain – The area between the coast and the Brooks Range. This area is sometimes split into coastal areas (lagoons, barrier islands, and Beaufort Sea) and inland areas (uplands near the foothills of the Brooks Range). Brooks Range – The mountains, valleys, and foothills north and south of the Continental Divide. South Side – The foothills, taiga, and boreal forest south of the Brooks Range. Status . Permanent Resident – Present throughout the year and breeds in the area. Summer Resident – Only present from May to September. Migrant – Travels through on the way to wintering or breeding areas. Breeder – Documented as a breeding species. Visitor – Present as a non-breeding species. * – Not documented. Abundance . Abundant – Very numerous in suitable habitats. Common – Very likely to be seen or heard in suitable habitats. Fairly Common – Numerous but not always present in suitable habitats. Uncommon – Occurs regularly but not always observed because of lower abundance or secretive behaviors.