Trapidil Inhibits Human Mesangiai Cell Proliferation: Effect on PDGF Β
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
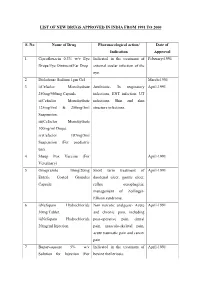
List of New Drugs Approved in India from 1991 to 2000
LIST OF NEW DRUGS APPROVED IN INDIA FROM 1991 TO 2000 S. No Name of Drug Pharmacological action/ Date of Indication Approval 1 Ciprofloxacin 0.3% w/v Eye Indicated in the treatment of February-1991 Drops/Eye Ointment/Ear Drop external ocular infection of the eye. 2 Diclofenac Sodium 1gm Gel March-1991 3 i)Cefaclor Monohydrate Antibiotic- In respiratory April-1991 250mg/500mg Capsule. infections, ENT infection, UT ii)Cefaclor Monohydrate infections, Skin and skin 125mg/5ml & 250mg/5ml structure infections. Suspension. iii)Cefaclor Monohydrate 100mg/ml Drops. iv)Cefaclor 187mg/5ml Suspension (For paediatric use). 4 Sheep Pox Vaccine (For April-1991 Veterinary) 5 Omeprazole 10mg/20mg Short term treatment of April-1991 Enteric Coated Granules duodenal ulcer, gastric ulcer, Capsule reflux oesophagitis, management of Zollinger- Ellison syndrome. 6 i)Nefopam Hydrochloride Non narcotic analgesic- Acute April-1991 30mg Tablet. and chronic pain, including ii)Nefopam Hydrochloride post-operative pain, dental 20mg/ml Injection. pain, musculo-skeletal pain, acute traumatic pain and cancer pain. 7 Buparvaquone 5% w/v Indicated in the treatment of April-1991 Solution for Injection (For bovine theileriosis. Veterinary) 8 i)Kitotifen Fumerate 1mg Anti asthmatic drug- Indicated May-1991 Tablet in prophylactic treatment of ii)Kitotifen Fumerate Syrup bronchial asthma, symptomatic iii)Ketotifen Fumerate Nasal improvement of allergic Drops conditions including rhinitis and conjunctivitis. 9 i)Pefloxacin Mesylate Antibacterial- In the treatment May-1991 Dihydrate 400mg Film Coated of severe infection in adults Tablet caused by sensitive ii)Pefloxacin Mesylate microorganism (gram -ve Dihydrate 400mg/5ml Injection pathogens and staphylococci). iii)Pefloxacin Mesylate Dihydrate 400mg I.V Bottles of 100ml/200ml 10 Ofloxacin 100mg/50ml & Indicated in RTI, UTI, May-1991 200mg/100ml vial Infusion gynaecological infection, skin/soft lesion infection. -

18512582.Pdf
European Heart Journal (2001) 22, 1938–1947 doi:10.1053/euhj.2001.2627, available online at http://www.idealibrary.com on The TRAPIST Study A multicentre randomized placebo controlled clinical trial of trapidil for prevention of restenosis after coronary stenting, measured by 3-D intravascular ultrasound P. W. Serruys1, D. P. Foley1, M. Pieper2, J. A. Kleijne3 and P. J. de Feyter1 on behalf of the TRAPIST investigators 1Department of Interventional Cardiology, Thoraxcenter, Erasmus Medical Center, Rotterdam, The Netherlands; 2Heartcenter Bodensee, Kreuzlingen, Switzerland; and 3Cardialysis, Rotterdam, The Netherlands Background Studies have reported benefit of oral therapy significant difference between trapidil and placebo-treated with the phosphodiesterase inhibitor, trapidil, in reducing patients regarding in-stent neointimal volume (108·6 restenosis after coronary angioplasty. Coronary stenting is 95·6 mm3 vs 93·379·1 mm3; P=0·16) or % obstruction associated with improved late outcome compared with volume (3818% vs 3621%; P=0·32), in angiographic balloon angioplasty, but significant neointimal hyperplasia minimal luminal diameter at follow-up (1·630·61 mm vs still occurs in a considerable proportion of patients. The 1·740·69 mm; P=0·17), restenosis rate (31% vs 24%; aim of this study was to investigate the safety and efficacy P=0·24), cumulative incidence of major adverse cardiac of trapidil 200 mg in preventing in-stent restenosis. events at 7 months (22% vs 20%; P=0·71) or anginal complaints (30% vs 24%; P=0·29). Methods Patients with a single native coronary lesion requiring revascularization were randomized to placebo or Conclusion Oral trapidil 600 mg daily for 6 months did trapidil at least 1 h before, and continuing for 6 months not reduce in-stent hyperplasia or improve clinical outcome after, successful implantation of a coronary Wallstent. -

(12) United States Patent (10) Patent No.: US 7.803,838 B2 Davis Et Al
USOO7803838B2 (12) United States Patent (10) Patent No.: US 7.803,838 B2 Davis et al. (45) Date of Patent: Sep. 28, 2010 (54) COMPOSITIONS COMPRISING NEBIVOLOL 2002fO169134 A1 11/2002 Davis 2002/0177586 A1 11/2002 Egan et al. (75) Inventors: Eric Davis, Morgantown, WV (US); 2002/0183305 A1 12/2002 Davis et al. John O'Donnell, Morgantown, WV 2002/0183317 A1 12/2002 Wagle et al. (US); Peter Bottini, Morgantown, WV 2002/0183365 A1 12/2002 Wagle et al. (US) 2002/0192203 A1 12, 2002 Cho 2003, OOO4194 A1 1, 2003 Gall (73) Assignee: Forest Laboratories Holdings Limited 2003, OO13699 A1 1/2003 Davis et al. (BM) 2003/0027820 A1 2, 2003 Gall (*) Notice: Subject to any disclaimer, the term of this 2003.0053981 A1 3/2003 Davis et al. patent is extended or adjusted under 35 2003, OO60489 A1 3/2003 Buckingham U.S.C. 154(b) by 455 days. 2003, OO69221 A1 4/2003 Kosoglou et al. 2003/0078190 A1* 4/2003 Weinberg ...................... 514f1 (21) Appl. No.: 11/141,235 2003/0078517 A1 4/2003 Kensey 2003/01 19428 A1 6/2003 Davis et al. (22) Filed: May 31, 2005 2003/01 19757 A1 6/2003 Davis 2003/01 19796 A1 6/2003 Strony (65) Prior Publication Data 2003.01.19808 A1 6/2003 LeBeaut et al. US 2005/027281.0 A1 Dec. 8, 2005 2003.01.19809 A1 6/2003 Davis 2003,0162824 A1 8, 2003 Krul Related U.S. Application Data 2003/0175344 A1 9, 2003 Waldet al. (60) Provisional application No. 60/577,423, filed on Jun. -

Antithrombotic Treatment After Stroke Due to Intracerebral Haemorrhage (Review)
Cochrane Database of Systematic Reviews Antithrombotic treatment after stroke due to intracerebral haemorrhage (Review) Perry LA, Berge E, Bowditch J, Forfang E, Rønning OM, Hankey GJ, Villanueva E, Al-Shahi Salman R Perry LA, Berge E, Bowditch J, Forfang E, Rønning OM, Hankey GJ, Villanueva E, Al-Shahi Salman R. Antithrombotic treatment after stroke due to intracerebral haemorrhage. Cochrane Database of Systematic Reviews 2017, Issue 5. Art. No.: CD012144. DOI: 10.1002/14651858.CD012144.pub2. www.cochranelibrary.com Antithrombotic treatment after stroke due to intracerebral haemorrhage (Review) Copyright © 2017 The Cochrane Collaboration. Published by John Wiley & Sons, Ltd. TABLE OF CONTENTS HEADER....................................... 1 ABSTRACT ...................................... 1 PLAINLANGUAGESUMMARY . 2 SUMMARY OF FINDINGS FOR THE MAIN COMPARISON . ..... 3 BACKGROUND .................................... 5 OBJECTIVES ..................................... 5 METHODS ...................................... 6 RESULTS....................................... 8 Figure1. ..................................... 9 Figure2. ..................................... 11 Figure3. ..................................... 12 DISCUSSION ..................................... 14 AUTHORS’CONCLUSIONS . 15 ACKNOWLEDGEMENTS . 15 REFERENCES ..................................... 15 CHARACTERISTICSOFSTUDIES . 18 DATAANDANALYSES. 31 Analysis 1.2. Comparison 1 Short-term antithrombotic treatment, Outcome 2 Death. 31 Analysis 1.6. Comparison 1 Short-term antithrombotic -

The Use of Stems in the Selection of International Nonproprietary Names (INN) for Pharmaceutical Substances
WHO/PSM/QSM/2006.3 The use of stems in the selection of International Nonproprietary Names (INN) for pharmaceutical substances 2006 Programme on International Nonproprietary Names (INN) Quality Assurance and Safety: Medicines Medicines Policy and Standards The use of stems in the selection of International Nonproprietary Names (INN) for pharmaceutical substances FORMER DOCUMENT NUMBER: WHO/PHARM S/NOM 15 © World Health Organization 2006 All rights reserved. Publications of the World Health Organization can be obtained from WHO Press, World Health Organization, 20 Avenue Appia, 1211 Geneva 27, Switzerland (tel.: +41 22 791 3264; fax: +41 22 791 4857; e-mail: [email protected]). Requests for permission to reproduce or translate WHO publications – whether for sale or for noncommercial distribution – should be addressed to WHO Press, at the above address (fax: +41 22 791 4806; e-mail: [email protected]). The designations employed and the presentation of the material in this publication do not imply the expression of any opinion whatsoever on the part of the World Health Organization concerning the legal status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries. Dotted lines on maps represent approximate border lines for which there may not yet be full agreement. The mention of specific companies or of certain manufacturers’ products does not imply that they are endorsed or recommended by the World Health Organization in preference to others of a similar nature that are not mentioned. Errors and omissions excepted, the names of proprietary products are distinguished by initial capital letters. -
![Ehealth DSI [Ehdsi V2.2.2-OR] Ehealth DSI – Master Value Set](https://docslib.b-cdn.net/cover/8870/ehealth-dsi-ehdsi-v2-2-2-or-ehealth-dsi-master-value-set-1028870.webp)
Ehealth DSI [Ehdsi V2.2.2-OR] Ehealth DSI – Master Value Set
MTC eHealth DSI [eHDSI v2.2.2-OR] eHealth DSI – Master Value Set Catalogue Responsible : eHDSI Solution Provider PublishDate : Wed Nov 08 16:16:10 CET 2017 © eHealth DSI eHDSI Solution Provider v2.2.2-OR Wed Nov 08 16:16:10 CET 2017 Page 1 of 490 MTC Table of Contents epSOSActiveIngredient 4 epSOSAdministrativeGender 148 epSOSAdverseEventType 149 epSOSAllergenNoDrugs 150 epSOSBloodGroup 155 epSOSBloodPressure 156 epSOSCodeNoMedication 157 epSOSCodeProb 158 epSOSConfidentiality 159 epSOSCountry 160 epSOSDisplayLabel 167 epSOSDocumentCode 170 epSOSDoseForm 171 epSOSHealthcareProfessionalRoles 184 epSOSIllnessesandDisorders 186 epSOSLanguage 448 epSOSMedicalDevices 458 epSOSNullFavor 461 epSOSPackage 462 © eHealth DSI eHDSI Solution Provider v2.2.2-OR Wed Nov 08 16:16:10 CET 2017 Page 2 of 490 MTC epSOSPersonalRelationship 464 epSOSPregnancyInformation 466 epSOSProcedures 467 epSOSReactionAllergy 470 epSOSResolutionOutcome 472 epSOSRoleClass 473 epSOSRouteofAdministration 474 epSOSSections 477 epSOSSeverity 478 epSOSSocialHistory 479 epSOSStatusCode 480 epSOSSubstitutionCode 481 epSOSTelecomAddress 482 epSOSTimingEvent 483 epSOSUnits 484 epSOSUnknownInformation 487 epSOSVaccine 488 © eHealth DSI eHDSI Solution Provider v2.2.2-OR Wed Nov 08 16:16:10 CET 2017 Page 3 of 490 MTC epSOSActiveIngredient epSOSActiveIngredient Value Set ID 1.3.6.1.4.1.12559.11.10.1.3.1.42.24 TRANSLATIONS Code System ID Code System Version Concept Code Description (FSN) 2.16.840.1.113883.6.73 2017-01 A ALIMENTARY TRACT AND METABOLISM 2.16.840.1.113883.6.73 2017-01 -

Search Strategy, Baseline Risk Coronavirus and Prophylaxis For
Supplement 7: Search Strategy, Baseline Risk Coronavirus and prophylaxis for thrombotic events Search narrative, 23 July 2020 Bibliographic databases: EMBASE (1974 to, 22 July 2020) Epistemonikos COVID-19 Evidence (21 July 2020) (includes records from Cochrane Database of Systematic Reviews, Pubmed, EMBASE, CINAHL, PsycINFO, LILACS, Database of reviews of Effects, The Campbell Collaboration online library, JBI Database of Systematic Reviews and Implementation Reports, EPPI-Centre Evidence Library) MEDLINE (1946 to, 23 July 2020) SCOPUS (19 July 2020) WHO Global Research Database (COVID-19) (20 July 2020) Trial/study Databases: Cochrane COVID-19 study register (23 July 2020) (includes records from ClinicalTrials.gov, WHO ICTRN, and PubMed) CYTEL map of ongoing [COVID-19] clinical trials (20 July 2020) (WHO International Clinical Trials Registry Platform, European Clinical Trials Registry, clinicaltrials.gov, Chinese Clinical Trial Registry, German Clinical Trials registry, Japan Primary Registries Network, Iranian Clinical Trial Registry, and Australian New Zealand Clinical Trials Registry) Results were downloaded from sources and deduplicated in EndNote. Results from BioRxiV, ChemRxiV, MedRxiV (preprints indexed in Medline and Epistemonikos), or protocols for clinical trials were removed as they were beyond the scope of the search. COCHRANE COVID-19 (https://covid-19.cochrane.org/) Search 1: anticoagula* or anti-coagula* or coagulation or thromb* or antithromb* or antiplatelet* or heparin or LMWH or aspirin or embol* or phlebothrombos* -

Total Thrombus-Formation Analysis System (T-TAS): Clinical Application of Quantitative Analysis of Thrombus Formation in Cardiovascular Disease
Published online: 2019-07-22 THIEME 1554 Theme Issue Article Total Thrombus-Formation Analysis System (T-TAS): Clinical Application of Quantitative Analysis of Thrombus Formation in Cardiovascular Disease Koichi Kaikita1 Kazuya Hosokawa2 Jeffrey R. Dahlen3 Kenichi Tsujita1 1 Department of Cardiovascular Medicine, Graduate School of Medical Address for correspondence Koichi Kaikita, MD, PhD, Department of Sciences, Kumamoto University, Kumamoto, Japan Cardiovascular Medicine, Graduate School of Medical Sciences, 2 Research Institute, Fujimori Kogyo Co., Yokohama, Kanagawa, Japan Kumamoto University, 1-1-1, Honjo, Chuo-ku, Kumamoto, 860-8556, 3 Hikari Dx, San Diego, California, United States Japan (e-mail: [email protected]). Thromb Haemost 2019;119:1554–1562. Abstract Various antithrombotic agents are clinically used to inhibit the cascade of arterial or venous thrombosis in cardiovascular diseases. Dual antiplatelet therapy with aspirin and P2Y12 inhibitors is prescribed in patients with coronary artery disease (CAD) undergoing percutaneous coronary intervention (PCI). Direct oral anticoagulants (DOACs) are widely used for the prevention or treatment of thromboembolism in patients with atrial fibrillation (AF) and venous thromboembolism. However, there has been no definitive tool to simultaneously monitor the antithrombotic effects of these drugs. The Total Thrombus-Formation Analysis System (T-TAS), a microchip-based flow chamber system that mimics in vivo conditions for evaluating whole blood thrombo- genicity, was developed for the quantitative analysis of thrombus formation in whole blood specimens. The utility of T-TAS has been evaluated in CAD patients treated with antiplatelet therapies. The T-TAS PL chip area under the flow pressure curve (AUC) accurately assesses primary hemostasis and is sensitive to the therapeutic effects of Keywords various antiplatelet therapies. -

(12) United States Patent (10) Patent No.: US 7,022,521 B2 Hanson (45) Date of Patent: * Apr
US007022521B2 (12) United States Patent (10) Patent No.: US 7,022,521 B2 Hanson (45) Date of Patent: * Apr. 4, 2006 (54) METHODS AND COMPOSITIONS FOR 5.440,020 A 8, 1995 Coller TREATING PLATELET RELATED 5,498.599 A 3, 1996 Choi et al. DSORDERS USING MIPL PATHWAY 5,506,107 A 4/1996 Cunningham et al. INHIBITORY AGENTS 5,572,944 A 11/1996 Slikkers et al. 5,620,960 A 4/1997 Arnold et al. 5,756,083. A 5, 1998 Elliott (75) Inventor: Stephen R. Hanson, Beaverton, OR 5,789,539 A 8/1998 Daly et al. (US) 5,830,647 A 11/1998 Eaton et al. 5,869,451 A 2f1999 Dower et al. (73) Assignee: Emory University, Atlanta, GA (US) 5,932,546 A 8, 1999 Barrett et al. 6,008,232 A 12/1999 Lakshmanan (*) Notice: Subject to any disclaimer, the term of this 6,103,740 A 8/2000 Lakshmanan patent is extended or adjusted under 35 6,150,120 A 11/2000 Spivak et al. U.S.C. 154(b) by 0 days. 6,376,242 B1 4/2002 Hanson 6,585,995 B1 7/2003 Hanson This patent is Subject to a terminal dis 2004/0087486 A1 5, 2004 Hanson claimer. FOREIGN PATENT DOCUMENTS (21) Appl. No.: 10/602,293 EP O 904 T83 3, 1999 WO WO 96,25498 8, 1996 (22) Filed: Jun. 24, 2003 WO WO 96/4O750 12/1996 WO WO 97.28190 8, 1997 (65) Prior Publication Data WO WO 99,08524 2, 1999 WO WO 99,08525 2, 1999 US 2004/OO87035 A1 May 6, 2004 WO WO 99.10494 4f1999 Related U.S. -
![Gann, 75, 284-291; March, 1984]](https://docslib.b-cdn.net/cover/9982/gann-75-284-291-march-1984-2529982.webp)
Gann, 75, 284-291; March, 1984]
[Gann, 75, 284-291; March, 1984] EFFECTS OF ANTIPLATELET AGENTS ON PULMONARY METASTASES Hiroyasu BANDO,Takashi YAMASHITAand Eiro TSUBURA ThirdDepartment of InternalMedicine, The Universityof Tokushima,School of Medicine, Kuramoto-cho3-chome,T okushima770 The role of platelets in cancer metastasis was studied by investigating the effects of the antiplatelet agents ticlopidine, diltiazem, dipyridamole and trapidil on arti ficialand spontaneous pulmonary metastases in mice. These agents were tested at their optimal inhibitory doses on adenosine diphosphate-induced platelet aggrega tion;namely, 100mg/kg for ticlopidine, 2mg/kg for diltiazem, 180mg/kg for trapidil and 60mg/kg for dipyridamole. At these doses, trapidil caused moderate inhibition of thrombin-induced platelet aggregation in mice, but the other agents had only slight effects. Artificial pulmonary metastasis was produced by inoculation of Lewis lung carcinoma (LLC) or B16 melanoma (B16) cells into C57BL/6 mice . For induction of spontaneous pulmonary metastases, these tumor cells were implanted subcutane ouslyinto the footpads of mice. The resulting primary tumors of LLC and B16 were removed 9-10 and 17 days later, respectively. Artificial pulmonary metastases were inhibited significantly by all the antiplatelet agents tested . Spontaneous pulmonary metastases were markedly reduced only when these agents were given after removal of the primary tumor. The role of platelets is discussed with respect to thrombus formation in the lodgement of tumor cells and the participation of platelet -derived growth factor in the growth of metastatic foci. Key words: Blood-borne metastasis-Antiplatelet agents-B16 melanoma - Lewis lung carcinoma-Adenosine diphosphate-induced platelet aggregation The outcome of hematogenous metastasis To clarify the role of platelets in cancer depends on the interaction of tumor cells metastasis, we studied the effects of various with various host factors in the metastatic antiplatelet agents (platelet aggregation in process. -

2000 Dialysis of Drugs
2000 Dialysis of Drugs PROVIDED AS AN EDUCATIONAL SERVICE BY AMGEN INC. I 2000 DIAL Dialysis of Drugs YSIS OF DRUGS Curtis A. Johnson, PharmD Member, Board of Directors Nephrology Pharmacy Associates Ann Arbor, Michigan and Professor of Pharmacy and Medicine University of Wisconsin-Madison Madison, Wisconsin William D. Simmons, RPh Senior Clinical Pharmacist Department of Pharmacy University of Wisconsin Hospital and Clinics Madison, Wisconsin SEE DISCLAIMER REGARDING USE OF THIS POCKET BOOK DISCLAIMER—These Dialysis of Drugs guidelines are offered as a general summary of information for pharmacists and other medical professionals. Inappropriate administration of drugs may involve serious medical risks to the patient which can only be identified by medical professionals. Depending on the circumstances, the risks can be serious and can include severe injury, including death. These guidelines cannot identify medical risks specific to an individual patient or recommend patient treatment. These guidelines are not to be used as a substitute for professional training. The absence of typographical errors is not guaranteed. Use of these guidelines indicates acknowledgement that neither Nephrology Pharmacy Associates, Inc. nor Amgen Inc. will be responsible for any loss or injury, including death, sustained in connection with or as a result of the use of these guidelines. Readers should consult the complete information available in the package insert for each agent indicated before prescribing medications. Guides such as this one can only draw from information available as of the date of publication. Neither Nephrology Pharmacy Associates, Inc. nor Amgen Inc. is under any obligation to update information contained herein. Future medical advances or product information may affect or change the information provided. -

Mitigating the Inhibition of Human Bile Salt Export Pump by Drugs
DMD Fast Forward. Published on September 7, 2012 as DOI: 10.1124/dmd.112.047068 DMD FastThis Forward. article has not Published been copyedited on andSeptember formatted. The 7, final 2012 version as doi:10.1124/dmd.112.047068may differ from this version. DMD #47968 Mitigating the inhibition of human Bile Salt Export Pump by drugs: opportunities provided by physicochemical property modulation, in-silico modeling and structural modification Daniel J. Warner, Hongming Chen, Louis-David Cantin, J. Gerry Kenna, Simone Stahl, Clare L. Walker, Tobias Noeske. Department of Medicinal Chemistry, AstraZeneca R&D Montreal, Montreal, Quebec, H4S Downloaded from 1Z9, Canada (DJW, LDC) Computational Sciences, Discovery Sciences, AstraZeneca R&D Mölndal, Pepparedsleden dmd.aspetjournals.org 1, Mölndal 43183, Sweden (HC) Molecular Toxicology, Global Safety Assessment, AstraZeneca, Alderley Park, Macclesfield, Cheshire, SK10 4TG, UK (JGK, SS, CLW) Global Safety Assessment, AstraZeneca R&D Mölndal, Pepparedsleden 1, Mölndal 43183, at ASPET Journals on October 10, 2021 Sweden (TN) 1 Copyright 2012 by the American Society for Pharmacology and Experimental Therapeutics. DMD Fast Forward. Published on September 7, 2012 as DOI: 10.1124/dmd.112.047068 This article has not been copyedited and formatted. The final version may differ from this version. DMD #47968 Inhibition of the human Bile Salt Export Pump by drugs. Corresponding author: Tobias Noeske Global Safety Assessment AstraZeneca R&D Mölndal S-431 83 Mölndal, Sweden Phone: +46-31-7064002 Mobile: +46-727-158344