Pervasive Use of Artificial Habitats by Shorebirds in the Asia-Pacific
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Aquatic Invertebrates and Waterbirds of Wetlands and Rivers of the Southern Carnarvon Basin, Western Australia
DOI: 10.18195/issn.0313-122x.61.2000.217-265 Records of the Western Australian Museum Supplement No. 61: 217-265 (2000). Aquatic invertebrates and waterbirds of wetlands and rivers of the southern Carnarvon Basin, Western Australia 3 3 S.A. Halsel, R.J. ShieF, A.W. Storey, D.H.D. Edward , I. Lansburyt, D.J. Cale and M.S. HarveyS 1 Department of Conservation and Land Management, Wildlife Research Centre, PO Box 51, Wanneroo, Western Australia 6946, Australia 2CRC for Freshwater Ecology, Murray-Darling Freshwater Research Centre, PO Box 921, Albury, New South Wales 2640, Australia 3 Department of Zoology, The University of Western Australia, Nedlands, Western Australia 6907, Australia 4 Hope Entomological Collections, Oxford University Museum, Parks Road, Oxford OXl 3PW, United Kingdom 5 Department of Terrestrial Invertebrates, Western Australian Museum, Francis Street, Perth, Western Australia 6000, Australia Abstract - Fifty-six sites, representing 53 wetlands, were surveyed in the southern Carnarvon Basin in 1994 and 1995 with the aim of documenting the waterbird and aquatic invertebrate fauna of the region. Most sites were surveyed in both winter and summer, although some contained water only one occasion. Altogether 57 waterbird species were recorded, with 29 292 waterbirds of 25 species on Lake MacLeod in October 1994. River pools were shown to be relatively important for waterbirds, while many freshwater claypans were little used. At least 492 species of aquatic invertebrate were collected. The invertebrate fauna was characterized by the low frequency with which taxa occurred: a third of the species were collected at a single site on only one occasion. -

Disaggregation of Bird Families Listed on Cms Appendix Ii
Convention on the Conservation of Migratory Species of Wild Animals 2nd Meeting of the Sessional Committee of the CMS Scientific Council (ScC-SC2) Bonn, Germany, 10 – 14 July 2017 UNEP/CMS/ScC-SC2/Inf.3 DISAGGREGATION OF BIRD FAMILIES LISTED ON CMS APPENDIX II (Prepared by the Appointed Councillors for Birds) Summary: The first meeting of the Sessional Committee of the Scientific Council identified the adoption of a new standard reference for avian taxonomy as an opportunity to disaggregate the higher-level taxa listed on Appendix II and to identify those that are considered to be migratory species and that have an unfavourable conservation status. The current paper presents an initial analysis of the higher-level disaggregation using the Handbook of the Birds of the World/BirdLife International Illustrated Checklist of the Birds of the World Volumes 1 and 2 taxonomy, and identifies the challenges in completing the analysis to identify all of the migratory species and the corresponding Range States. The document has been prepared by the COP Appointed Scientific Councilors for Birds. This is a supplementary paper to COP document UNEP/CMS/COP12/Doc.25.3 on Taxonomy and Nomenclature UNEP/CMS/ScC-Sc2/Inf.3 DISAGGREGATION OF BIRD FAMILIES LISTED ON CMS APPENDIX II 1. Through Resolution 11.19, the Conference of Parties adopted as the standard reference for bird taxonomy and nomenclature for Non-Passerine species the Handbook of the Birds of the World/BirdLife International Illustrated Checklist of the Birds of the World, Volume 1: Non-Passerines, by Josep del Hoyo and Nigel J. Collar (2014); 2. -

A Guide to the Birds of Barrow Island
A Guide to the Birds of Barrow Island Operated by Chevron Australia This document has been printed by a Sustainable Green Printer on stock that is certified carbon in joint venture with neutral and is Forestry Stewardship Council (FSC) mix certified, ensuring fibres are sourced from certified and well managed forests. The stock 55% recycled (30% pre consumer, 25% post- Cert no. L2/0011.2010 consumer) and has an ISO 14001 Environmental Certification. ISBN 978-0-9871120-1-9 Gorgon Project Osaka Gas | Tokyo Gas | Chubu Electric Power Chevron’s Policy on Working in Sensitive Areas Protecting the safety and health of people and the environment is a Chevron core value. About the Authors Therefore, we: • Strive to design our facilities and conduct our operations to avoid adverse impacts to human health and to operate in an environmentally sound, reliable and Dr Dorian Moro efficient manner. • Conduct our operations responsibly in all areas, including environments with sensitive Dorian Moro works for Chevron Australia as the Terrestrial Ecologist biological characteristics. in the Australasia Strategic Business Unit. His Bachelor of Science Chevron strives to avoid or reduce significant risks and impacts our projects and (Hons) studies at La Trobe University (Victoria), focused on small operations may pose to sensitive species, habitats and ecosystems. This means that we: mammal communities in coastal areas of Victoria. His PhD (University • Integrate biodiversity into our business decision-making and management through our of Western Australia) -

Habitat Types
Habitat Types The following section features ten predominant habitat types on the West Coast of the Eyre Peninsula, South Australia. It provides a description of each habitat type and the native plant and fauna species that commonly occur there. The fauna species lists in this section are not limited to the species included in this publication and include other coastal fauna species. Fauna species included in this publication are printed in bold. Information is also provided on specific threats and reference sites for each habitat type. The habitat types presented are generally either characteristic of high-energy exposed coastline or low-energy sheltered coastline. Open sandy beaches, non-vegetated dunefields, coastal cliffs and cliff tops are all typically found along high energy, exposed coastline, while mangroves, sand flats and saltmarsh/samphire are characteristic of low energy, sheltered coastline. Habitat Types Coastal Dune Shrublands NATURAL DISTRIBUTION shrublands of larger vegetation occur on more stable dunes and Found throughout the coastal environment, from low beachfront cliff-top dunes with deep stable sand. Most large dune shrublands locations to elevated clifftops, wherever sand can accumulate. will be composed of a mosaic of transitional vegetation patches ranging from bare sand to dense shrub cover. DESCRIPTION This habitat type is associated with sandy coastal dunes occurring The understory generally consists of moderate to high diversity of along exposed and sometimes more sheltered coastline. Dunes are low shrubs, sedges and groundcovers. Understory diversity is often created by the deposition of dry sand particles from the beach by driven by the position and aspect of the dune slope. -

Biology and Conservation of the Unique and Diverse Halophilic Macroinvertebrates of Australian Salt Lakes
CSIRO PUBLISHING Marine and Freshwater Research Corrigendum https://doi.org/10.1071/MF21088_CO Biology and conservation of the unique and diverse halophilic macroinvertebrates of Australian salt lakes Angus D’Arcy Lawrie, Jennifer Chaplin and Adrian Pinder Marine and Freshwater Research. [Published online 2 July 2021]. https://doi.org/10.1071/MF21088 The authors of the above-mentioned paper regret to inform readers that there were errors published in the systematics of one of the taxa in the manuscript. The list of groups in the Cladocera section (on p. F) was published as below: The bulk of Cladocera that occur in inland waters in Australia are restricted to fresh water, but three groups have representatives in salt lakes. These groups comprise: (1) six species of Daphniopsis (or Daphnia; see below); (2) two species of Daphnia (Daphnia salinifera Hebert and Daphnia neosalinifera Hebert) from the Daphnia carinata (King) subgenus; and (3) three species of chydorid: Moina baylyi Forro´, Moina mongolica Daday and Extremalona timmsi Sinev & Shiel. This text should have been as below (changes underlined): The bulk of Cladocera that occur in inland waters in Australia are restricted to fresh water, but four groups have representatives in salt lakes. These groups comprise: (1) six species of Daphniopsis (or Daphnia; see below); (2) two species of Daphnia (Daphnia salinifera Hebert and Daphnia neosalinifera Hebert) from the Daphnia carinata (King) subgenus; (3) two Moina species (Moina baylyi Forro´ and Moina mongolica Daday); and (4) one species of chydorid (Extremalona timmsi Sinev & Shiel). Furthermore, the title of the Chydorids section should have been titled Moinids and chydorids. -

The Maintenance and Reproductive Behaviour of Black Stilts
Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and private study only. The thesis may not be reproduced elsewhere without the pennission of the Author. The Maintenance and Reproductive Behaviour of Black Stilts (Himantopus novaezealandiae) in Captivity, and Implications for the Management of this Rare Species. A Thesis Presented in Partial Fulfilment of the Requirements for the Degree of Master of Science in Zoology at Massey University Christine Elva Margaret Reed February 1986 ii ABSTRACT In an effort to conserve New Zealand's rarest endemic wading species, the black stilt (Himantopus novaezealandiae), eggs were removed from the wild in October 1979 for establishment of a captive breeding population. Eight chicks fledged following artificial incubation and hand-rearing at the National Wildlife Centre near Masterton. At two years of age, these tentatively sexed stilts were formed into pairs and housed in large outdoor enclosures. I studied their behaviour from December 1982 until February 1986, aiming to i) collate an ethogram for the species under the restrictions of a captive environment ii) describe and quantify behavioural activity, especially that of reproductive behaviour and breeding biology iii) describe vocalisations and iv) on the basis of observed behaviour, examine captive breeding as a management option for black stilts. A repertoire of 38 context-specific behavioural patterns were observed throughout the year and a further 15 stereo-typed species-typical nest-building, copulatory, incubation and chick-rearing patterns occurred during the breeding season. Time-budget analysis of a focal pair of stilts showed foraging and immobility to be the predominant daily activities, peaking during the pre-nesting period. -

Preliminary Flora and Fauna Assessment - Penshurst Wind Farm Final.Doc
Preliminary Flora and Fauna Assessment - Penshurst Wind Farm Project: 09 - 055 Prepared for: RES Australia Ecology Australia Pty Ltd Flora and Fauna Consultants www.ecologyaustralia.com.au [email protected] 88B Station Street, Fairfield, Victoria, Australia 3078 Tel: (03) 9489 4191 Fax: (03) 9481 7679 © 2009 Ecology Australia Pty Ltd This publication is copyright. It may only be used in accordance with the agreed terms of the commission. Except as provided for by the Copyright Act 1968, no part of this publication may be reproduced, stored in a retrieval system, or transmitted in any form or by any means, without prior written permission of Ecology Australia Pty Ltd. Document information This is a controlled document. Details of the document ownership, location, distribution, status and revision history are listed below. All comments or requests for changes to content should be addressed to the document owner. Bioregion: Victorian Volcanic Plain Owner Ecology Australia Author Andrew McMahon and Ruth Marr J:\CURRENT PROJECTS\Penshurst Windfarm 09- Location 55\report\Preliminary Flora and Fauna Assessment - Penshurst Wind Farm Final.doc Distribution Simon Kerrison RES Australia Document History Status Changes By Date Draft 0.1 First Draft Andrew McMahon 22/7/09 and Ruth Marr Final Final Andrew McMahon 10/08/09 and Ruth Marr Final - ii Preliminary Flora and Fauna Assessment - Penshurst Wind Farm Contents Summary 1 1 Introduction 1 2 Study Area 3 3 Methods 5 3.1 Information Review 5 3.2 Preliminary site assessment 6 3.3 Liaison -

Dietary Breadth and Foraging Habitats of the White- Bellied Sea Eagle (Haliaeetus Leucogaster) on West Australian Islands and Coastal Sites
Dietary breadth and foraging habitats of the White- bellied Sea Eagle (Haliaeetus leucogaster) on West Australian islands and coastal sites. Presented to the Faculty of the Department of Environmental Science Murdoch University By Shannon Clohessy Bachelor of Science (Biological Sciences and Marine and Freshwater Management) Graduate Diploma of Science (Environmental Management) 2014 1 Declaration I declare that this thesis is a synthesis of my own research and has not been submitted as part of a tertiary qualification at any other institution. ……………………………………….. Shannon Clohessy 2014 2 Abstract This study looks at dietary preference of the Haliaeetus leucogaster in the Houtman Abrolhos and on coastal and near shore islands between Shark Bay and Jurien Bay. Prey species were identified through pellet dissection, which were collected from nests and feeding butcheries, along with prey remains and reference photographs. Data extracted from this process was compared against known prey types for this species. Potential foraging distances were calculated based on congeneric species data and feeding habits and used to calculate foraging habitat in the study sites and expected prey lists to compare against observed finds. Results were compared against similar studies on Haliaeetus leucogaster based in other parts of Australia. 3 Contents Figure list .................................................................................................................................. 6 Tables list ................................................................................................................................ -

Information Sheet on Ramsar Wetlands Categories Approved by Recommendation 4.7 of the Conference of the Contracting Parties
Information Sheet on Ramsar Wetlands Categories approved by Recommendation 4.7 of the Conference of the Contracting Parties. 1. Date this sheet was completed/updated: FOR OFFICE USE ONLY. May 1999 DD MM YY 2. Country: Australia Designation date Site Reference Number 3. Name of wetland: Western District Lakes, Victoria 4. Geographical coordinates: Latitude: 380 00' to 380 20'S; Longitude: 1430 07' to 1430 55'E 5. Altitude: Approximately 40 metres. 6. Area: 32,898 ha Note: This is a revised area figure based on GIS Mapping (1995) and does not represent any change to the Ramsar Site boundary. 7. Overview: The Western District Lakes, are important geomorphic features of a basaltic landscape. Water regimes vary both seasonally and annually so at any time the various lakes range from fresh to hypersaline. They support large numbers of mainly non-breeding waterbirds and are particularly important during periods of widespread drought and as moulting sites for some species. 8. Wetland Type: marine-coastal: A B C D E F G H I J K inland: L M N O P Q R Sp Ss Tp Ts U Va Vt W Xf Xp Y Zg Zk man-made: 1 2 3 4 5 6 7 8 9 9. Ramsar Criteria: 1a 1b 1c 1d 2a 2b 2c 2d 3a 3b 3c 4a 4b Please specify the most significant criterion applicable to the site: 10. Map of site included? Please tick yes -or- no.⌧ 11. Name and address of the compiler of this form: Simon Casanelia Parks Victoria 378 Cotham Road Kew VIC 3101 Australia Telephone 613 9816 1163 Facsimile 613 9816 9799 12. -
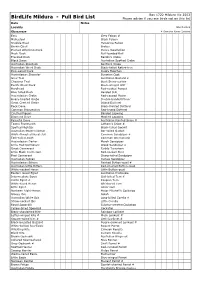
SBOC Full Bird List
Box 1722 Mildura Vic 3502 BirdLife Mildura - Full Bird List Please advise if you see birds not on this list Date Notes Locality March 2012 Observers # Denotes Rarer Species Emu Grey Falcon # Malleefowl Black Falcon Stubble Quail Peregrine Falcon Brown Quail Brolga Plumed Whistling-Duck Purple Swamphen Musk Duck Buff-banded Rail Freckled Duck Baillon's Crake Black Swan Australian Spotted Crake Australian Shelduck Spotless Crake Australian Wood Duck Black-tailed Native-hen Pink-eared Duck Dusky Moorhen Australasian Shoveler Eurasian Coot Grey Teal Australian Bustard # Chestnut Teal Bush Stone-curlew Pacific Black Duck Black-winged Stilt Hardhead Red-necked Avocet Blue-billed Duck Banded Stilt Australasian Grebe Red-capped Plover Hoary-headed Grebe Double-banded Plover Great Crested Grebe Inland Dotterel Rock Dove Black-fronted Dotterel Common Bronzewing Red-kneed Dotterel Crested Pigeon Banded Lapwing Diamond Dove Masked Lapwing Peaceful Dove Australian Painted Snipe # Tawny Frogmouth Latham's Snipe # Spotted Nightjar Black-tailed Godwit Australian Owlet-nightjar Bar-tailed Godwit White-throated Needletail Common Sandpiper # Fork-tailed Swift Common Greenshank Australasian Darter Marsh Sandpiper Little Pied Cormorant Wood Sandpiper # Great Cormorant Ruddy Turnstone Little Black Cormorant Red-necked Stint Pied Cormorant Sharp-tailed Sandpiper Australian Pelican Curlew Sandpiper Australasian Bittern Painted Button-quail # Australian Little Bittern Red-chested Button-quail White-necked Heron Little Button-quail Eastern Great Egret Australian -

Disease Risk Analysis for Birds at Ramsar Wetlands in Port Phillip Bay (Western Shoreline) and Bellarine Peninsula
Disease Risk Analysis for Birds at Ramsar Wetlands in Port Phillip Bay (Western Shoreline) and Bellarine Peninsula 11 - 12 March 2020 Disclaimer This Disease Risk Analysis (DRA) Worksheet has been developed by Auckland Zoo on behalf of the IUCN-SSC Conservation Planning Specialist Group (CPSG) to provide a structured template for the capture of outputs from wildlife disease risk analyses. CPSG encourages the sharing of completed DRAs using this template to assist other and progress the science of wildlife DRA, no responsibility is accepted for the validity of data, opinions or recommendations contained therein. Anyone who wishes to make their completed DRAs available to others can contact the CPSG office at [email protected] Wiethoelter A, Whiteley P, Gang R and Jakob-Hoff R (2020) Disease Risk Analysis for Birds at Ramsar Wetlands in Port Phillip Bay (Western Shoreline) and Bellarine Peninsula. IUCN SSC Conservation Planning Specialist Group: Apple Valley, MN. Cover photo ©: Anke Wiethoelter This report can be downloaded from the CPSG website: www.cpsg.org. 2 PPBBP Ramsar Wetland DRA, 2020 Table of Contents 37TList of Figures37T ............................................................................................................................................... 4 37TList of Tables37T ................................................................................................................................................ 5 37TACKNOWLEDMENTS37T ................................................................................................................................... -

The Wetlands of the Lake Carey Catchment, Northeast Goldfields of Western Australia, with Special Reference to Large Branchiopods
Journal of the Royal Society of Western Australia, 89: 175–183, 2006 The wetlands of the Lake Carey catchment, northeast Goldfields of Western Australia, with special reference to large branchiopods B V Timms1, B Datson2 & M Coleman2 1 School of Environmental and Life Sciences, University of Newcastle, Callaghan, NSW, Australia, 2265 2 actis Environmental Services, P.O. Box 176, Darlington, WA, Australia, 6070. Manuscript received June 2006; accepted July 2006 Abstract After significant rains, many areas of the Australian arid-zone contain numerous wetlands and support a diverse aquatic fauna; an example is Lake Carey and environs. Wetlands comprise 13.6% of the Carey catchment of nearly 6000 km2. Besides the large salina of Lake Carey (12.5% of catchment), there are many wetland types, including in order of total size: gypsum pans, samphire pans, salt pans, vegetated pans, red clay pans, creek pools and dams. Many of the smaller sites contain at least some water in late summer/autumn, and all are filled episodically to capacity at widely spaced intervals. Salinity is the most important factor distinguishing between sites, followed by turbidity and hydroperiod. Zooplankton and macroinvertebrates are mainly of widespread inland species, with coleopterans the most diverse, many odonatans, hemipterans and dipterans and a restricted fauna of oligochaetes, ephemeropterans, trichopterans and molluscs. The large branchiopod fauna is relatively diverse and comprises 12 anostracans in three genera (Branchinella, Streptocephalus, Parartemia), six clam shrimp in six genera: Lynceus (Laevicaudata) and Caenestheria, Caenestheriella, Eocyzicus, Limnadia, Limnadopsis (Spinicaudata)) and at least one notostracan (Triops). Almost all are widely distributed, though with many restrictions as to habitat type in the Carey wetlands.