A Tale of Two Tails Eric M. Jennings Janezic
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Genomic and Transcriptome Analysis Revealing an Oncogenic Functional Module in Meningiomas
Neurosurg Focus 35 (6):E3, 2013 ©AANS, 2013 Genomic and transcriptome analysis revealing an oncogenic functional module in meningiomas XIAO CHANG, PH.D.,1 LINGLING SHI, PH.D.,2 FAN GAO, PH.D.,1 JONATHAN RUssIN, M.D.,3 LIYUN ZENG, PH.D.,1 SHUHAN HE, B.S.,3 THOMAS C. CHEN, M.D.,3 STEVEN L. GIANNOTTA, M.D.,3 DANIEL J. WEISENBERGER, PH.D.,4 GAbrIEL ZADA, M.D.,3 KAI WANG, PH.D.,1,5,6 AND WIllIAM J. MAck, M.D.1,3 1Zilkha Neurogenetic Institute, Keck School of Medicine, University of Southern California, Los Angeles, California; 2GHM Institute of CNS Regeneration, Jinan University, Guangzhou, China; 3Department of Neurosurgery, Keck School of Medicine, University of Southern California, Los Angeles, California; 4USC Epigenome Center, Keck School of Medicine, University of Southern California, Los Angeles, California; 5Department of Psychiatry, Keck School of Medicine, University of Southern California, Los Angeles, California; and 6Division of Bioinformatics, Department of Preventive Medicine, Keck School of Medicine, University of Southern California, Los Angeles, California Object. Meningiomas are among the most common primary adult brain tumors. Although typically benign, roughly 2%–5% display malignant pathological features. The key molecular pathways involved in malignant trans- formation remain to be determined. Methods. Illumina expression microarrays were used to assess gene expression levels, and Illumina single- nucleotide polymorphism arrays were used to identify copy number variants in benign, atypical, and malignant me- ningiomas (19 tumors, including 4 malignant ones). The authors also reanalyzed 2 expression data sets generated on Affymetrix microarrays (n = 68, including 6 malignant ones; n = 56, including 3 malignant ones). -

Nº Ref Uniprot Proteína Péptidos Identificados Por MS/MS 1 P01024
Document downloaded from http://www.elsevier.es, day 26/09/2021. This copy is for personal use. Any transmission of this document by any media or format is strictly prohibited. Nº Ref Uniprot Proteína Péptidos identificados 1 P01024 CO3_HUMAN Complement C3 OS=Homo sapiens GN=C3 PE=1 SV=2 por 162MS/MS 2 P02751 FINC_HUMAN Fibronectin OS=Homo sapiens GN=FN1 PE=1 SV=4 131 3 P01023 A2MG_HUMAN Alpha-2-macroglobulin OS=Homo sapiens GN=A2M PE=1 SV=3 128 4 P0C0L4 CO4A_HUMAN Complement C4-A OS=Homo sapiens GN=C4A PE=1 SV=1 95 5 P04275 VWF_HUMAN von Willebrand factor OS=Homo sapiens GN=VWF PE=1 SV=4 81 6 P02675 FIBB_HUMAN Fibrinogen beta chain OS=Homo sapiens GN=FGB PE=1 SV=2 78 7 P01031 CO5_HUMAN Complement C5 OS=Homo sapiens GN=C5 PE=1 SV=4 66 8 P02768 ALBU_HUMAN Serum albumin OS=Homo sapiens GN=ALB PE=1 SV=2 66 9 P00450 CERU_HUMAN Ceruloplasmin OS=Homo sapiens GN=CP PE=1 SV=1 64 10 P02671 FIBA_HUMAN Fibrinogen alpha chain OS=Homo sapiens GN=FGA PE=1 SV=2 58 11 P08603 CFAH_HUMAN Complement factor H OS=Homo sapiens GN=CFH PE=1 SV=4 56 12 P02787 TRFE_HUMAN Serotransferrin OS=Homo sapiens GN=TF PE=1 SV=3 54 13 P00747 PLMN_HUMAN Plasminogen OS=Homo sapiens GN=PLG PE=1 SV=2 48 14 P02679 FIBG_HUMAN Fibrinogen gamma chain OS=Homo sapiens GN=FGG PE=1 SV=3 47 15 P01871 IGHM_HUMAN Ig mu chain C region OS=Homo sapiens GN=IGHM PE=1 SV=3 41 16 P04003 C4BPA_HUMAN C4b-binding protein alpha chain OS=Homo sapiens GN=C4BPA PE=1 SV=2 37 17 Q9Y6R7 FCGBP_HUMAN IgGFc-binding protein OS=Homo sapiens GN=FCGBP PE=1 SV=3 30 18 O43866 CD5L_HUMAN CD5 antigen-like OS=Homo -

Research2007herschkowitzetvolume Al
Open Access Research2007HerschkowitzetVolume al. 8, Issue 5, Article R76 Identification of conserved gene expression features between comment murine mammary carcinoma models and human breast tumors Jason I Herschkowitz¤*†, Karl Simin¤‡, Victor J Weigman§, Igor Mikaelian¶, Jerry Usary*¥, Zhiyuan Hu*¥, Karen E Rasmussen*¥, Laundette P Jones#, Shahin Assefnia#, Subhashini Chandrasekharan¥, Michael G Backlund†, Yuzhi Yin#, Andrey I Khramtsov**, Roy Bastein††, John Quackenbush††, Robert I Glazer#, Powel H Brown‡‡, Jeffrey E Green§§, Levy Kopelovich, reviews Priscilla A Furth#, Juan P Palazzo, Olufunmilayo I Olopade, Philip S Bernard††, Gary A Churchill¶, Terry Van Dyke*¥ and Charles M Perou*¥ Addresses: *Lineberger Comprehensive Cancer Center. †Curriculum in Genetics and Molecular Biology, University of North Carolina at Chapel Hill, Chapel Hill, NC 27599, USA. ‡Department of Cancer Biology, University of Massachusetts Medical School, Worcester, MA 01605, USA. reports §Department of Biology and Program in Bioinformatics and Computational Biology, University of North Carolina at Chapel Hill, Chapel Hill, NC 27599, USA. ¶The Jackson Laboratory, Bar Harbor, ME 04609, USA. ¥Department of Genetics, University of North Carolina at Chapel Hill, Chapel Hill, NC 27599, USA. #Department of Oncology, Lombardi Comprehensive Cancer Center, Georgetown University, Washington, DC 20057, USA. **Department of Pathology, University of Chicago, Chicago, IL 60637, USA. ††Department of Pathology, University of Utah School of Medicine, Salt Lake City, UT 84132, USA. ‡‡Baylor College of Medicine, Houston, TX 77030, USA. §§Transgenic Oncogenesis Group, Laboratory of Cancer Biology and Genetics. Chemoprevention Agent Development Research Group, National Cancer Institute, Bethesda, MD 20892, USA. Department of Pathology, Thomas Jefferson University, Philadelphia, PA 19107, USA. Section of Hematology/Oncology, Department of Medicine, Committees on Genetics and Cancer Biology, University of Chicago, Chicago, IL 60637, USA. -

Retrospective Analysis of the Two Sister Study Using Haplotype-Based Association Testing To
medRxiv preprint doi: https://doi.org/10.1101/2020.12.04.20244251; this version posted December 28, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted medRxiv a license to display the preprint in perpetuity. All rights reserved. No reuse allowed without permission. 1 Retrospective analysis of The Two Sister Study using haplotype-based association testing to 2 identify loci associated with early-onset breast cancer 3 James R. Gilbert, Ph.D. 1, James J. Cray, Ph.D. 2, Joseph E. Losee, M.D. 1, Gregory M. Cooper, 4 Ph.D. 1,3,4. 5 6 1. Department of Plastic Surgery, University of Pittsburgh/Children’s Hospital of Pittsburgh, 7 Pittsburgh, PA 15201. 8 2. Division of Anatomy, The Ohio State University College of Medicine, Columbus, OH 9 43210. 10 3. Department of Oral Biology, University of Pittsburgh/Children’s Hospital of Pittsburgh, 11 Pittsburgh, PA 15201. 12 4. Department of Bioengineering, University of Pittsburgh/Children’s Hospital of Pittsburgh, 13 Pittsburgh, PA 15201. 14 15 Email addresses: 16 [email protected] 17 [email protected] 18 [email protected] 19 [email protected] 20 21 Running Title: 22 23 Funding: The work described within this study was funded through the Children’s Fund of 24 Children’s Hospital of Pittsburgh of UPMC and through the Ross H. Musgrave Endowment 25 (J.E.L). 26 27 Conflicts of Interest: There are no conflicts of interest to disclose. 28 Corresponding Author: 29 Dr. Gregory M. -

Transposon Mutagenesis Identifies Genetic Drivers of Brafv600e Melanoma
ARTICLES Transposon mutagenesis identifies genetic drivers of BrafV600E melanoma Michael B Mann1,2, Michael A Black3, Devin J Jones1, Jerrold M Ward2,12, Christopher Chin Kuan Yew2,12, Justin Y Newberg1, Adam J Dupuy4, Alistair G Rust5,12, Marcus W Bosenberg6,7, Martin McMahon8,9, Cristin G Print10,11, Neal G Copeland1,2,13 & Nancy A Jenkins1,2,13 Although nearly half of human melanomas harbor oncogenic BRAFV600E mutations, the genetic events that cooperate with these mutations to drive melanogenesis are still largely unknown. Here we show that Sleeping Beauty (SB) transposon-mediated mutagenesis drives melanoma progression in BrafV600E mutant mice and identify 1,232 recurrently mutated candidate cancer genes (CCGs) from 70 SB-driven melanomas. CCGs are enriched in Wnt, PI3K, MAPK and netrin signaling pathway components and are more highly connected to one another than predicted by chance, indicating that SB targets cooperative genetic networks in melanoma. Human orthologs of >500 CCGs are enriched for mutations in human melanoma or showed statistically significant clinical associations between RNA abundance and survival of patients with metastatic melanoma. We also functionally validate CEP350 as a new tumor-suppressor gene in human melanoma. SB mutagenesis has thus helped to catalog the cooperative molecular mechanisms driving BRAFV600E melanoma and discover new genes with potential clinical importance in human melanoma. Substantial sun exposure and numerous genetic factors, including including BrafV600E, recapitulate the genetic and histological hallmarks skin type and family history, are the most important melanoma risk of human melanoma. In these models, increased MEK-ERK signaling factors. Familial melanoma, which accounts for <10% of cases, is asso- initiates clonal expansion of melanocytes, which is limited by oncogene- ciated with mutations in CDKN2A1, MITF2 and POT1 (refs. -

Multi-Omics of 34 Colorectal Cancer Cell Lines - a Resource for Biomedical Studies Kaja C
Berg et al. Molecular Cancer (2017) 16:116 DOI 10.1186/s12943-017-0691-y RESEARCH Open Access Multi-omics of 34 colorectal cancer cell lines - a resource for biomedical studies Kaja C. G. Berg1,2†, Peter W. Eide1,2†, Ina A. Eilertsen1,2, Bjarne Johannessen1,2,3, Jarle Bruun1,2, Stine A. Danielsen1,2,3, Merete Bjørnslett1,2, Leonardo A. Meza-Zepeda3,4, Mette Eknæs1,2, Guro E. Lind1,2, Ola Myklebost3,4,5, Rolf I. Skotheim1,2,3, Anita Sveen1,2,3 and Ragnhild A. Lothe1,2,3* Abstract Background: Colorectal cancer (CRC) cell lines are widely used pre-clinical model systems. Comprehensive insights into their molecular characteristics may improve model selection for biomedical studies. Methods: We have performed DNA, RNA and protein profiling of 34 cell lines, including (i) targeted deep sequencing (n = 612 genes) to detect single nucleotide variants and insertions/deletions; (ii) high resolution DNA copy number profiling; (iii) gene expression profiling at exon resolution; (iv) small RNA expression profiling by deep sequencing; and (v) protein expression analysis (n = 297 proteins) by reverse phase protein microarrays. Results: The cell lines were stratified according to the key molecular subtypes of CRC and data were integrated at two or more levels by computational analyses. We confirm that the frequencies and patterns of DNA aberrations are associated with genomic instability phenotypes and that the cell lines recapitulate the genomic profiles of primary carcinomas. Intrinsic expression subgroups are distinct from genomic subtypes, but consistent at the gene-, microRNA- and protein-level and dominated by two distinct clusters; colon-like cell lines characterized by expression of gastro-intestinal differentiation markers and undifferentiated cell lines showing upregulation of epithelial-mesenchymal transition and TGFβ signatures. -
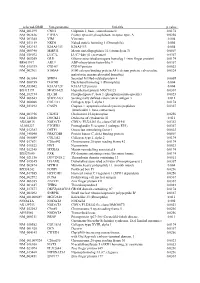
Supplementary Figures S1-S3
selected-GBID Uni-genename Uni-title p value NM_001299 CNN1 Calponin 1, basic, smooth muscle 0.0174 NM_002836 PTPRA Protein tyrosine phosphatase, receptor type, A 0.0256 NM_003380 VIM Vimentin 0.004 NM_033119 NKD1 Naked cuticle homolog 1 (Drosophila) 0.004 NM_052913 KIAA1913 KIAA1913 0.004 NM_005940 MMP11 Matrix metallopeptidase 11 (stromelysin 3) 0.0069 NM_018032 LUC7L LUC7-like (S. cerevisiae) 0.0367 NM_005269 GLI1 Glioma-associated oncogene homolog 1 (zinc finger protein) 0.0174 BE463997 ARL9 ADP-ribosylation factor-like 9 0.0367 NM_015939 CGI-09 CGI-09 protein 0.0023 NM_002961 S100A4 S100 calcium binding protein A4 (calcium protein, calvasculin, 0.0324 metastasin, murine placental homolog) NM_003014 SFRP4 Secreted frizzled-related protein 4 0.0005 NM_080759 DACH1 Dachshund homolog 1 (Drosophila) 0.004 NM_053042 KIAA1729 KIAA1729 protein 0.004 BX415194 MGC16121 Hypothetical protein MGC16121 0.0367 NM_182734 PLCB1 Phospholipase C, beta 1 (phosphoinositide-specific) 0.0023 NM_006643 SDCCAG3 Serologically defined colon cancer antigen 3 0.011 NM_000088 COL1A1 Collagen, type I, alpha 1 0.0174 NM_033292 CASP1 Caspase 1, apoptosis-related cysteine peptidase 0.0367 (interleukin 1, beta, convertase) NM_003956 CH25H Cholesterol 25-hydroxylase 0.0256 NM_144658 DOCK11 Dedicator of cytokinesis 11 0.011 AK024935 NODATA CDNA: FLJ21283 fis, clone COL01910 0.0363 AL050227 PTGER3 Prostaglandin E receptor 3 (subtype EP3) 0.0367 NM_012383 OSTF1 Osteoclast stimulating factor 1 0.0023 NM_145040 PRKCDBP Protein kinase C, delta binding protein 0.0069 NM_000089 -

Evaluation of Proteomic and Transcriptomic Biomarker Discovery
EVALUATION OF PROTEOMIC AND TRANSCRIPTOMIC BIOMARKER DISCOVERY TECHNOLOGIES IN OVARIAN CANCER. CLARE RITA ELIZABETH COVENEY A thesis submitted in partial fulfilment of the requirements of the Nottingham Trent University for the degree of Doctor of Philosophy October 2016 Copyright Statement “This work is the intellectual property of the author. You may copy up to 5% of this work for private study, or personal, non-commercial research. Any re-use of the information contained within this document should be fully referenced, quoting the author, title, university, degree level and pagination. Queries or requests for any other use, or if a more substantial copy is required, should be directed in the owner(s) of the Intellectual Property Rights.” Acknowledgments This work was funded by The John Lucille van Geest Foundation and undertaken at the John van Geest Cancer Research Centre, at Nottingham Trent University. I would like to extend my foremost gratitude to my supervisory team Professor Graham Ball, Dr David Boocock, Professor Robert Rees for their guidance, knowledge and advice throughout the course of this project. I would also like to show my appreciation of the hard work of Mr Ian Scott, Professor Bob Shaw and Dr Matharoo-Ball, Dr Suman Malhi and later Mr Viren Asher who alongside colleagues at The Nottingham University Medical School and Derby City General Hospital initiated the ovarian serum collection project that lead to this work. I also would like to acknowledge the work of Dr Suha Deen at Queen’s Medical Centre and Professor Andrew Green and Christopher Nolan of the Cancer & Stem Cells Division of the School of Medicine, University of Nottingham for support with the immunohistochemistry. -

Autocrine IFN Signaling Inducing Profibrotic Fibroblast Responses by a Synthetic TLR3 Ligand Mitigates
Downloaded from http://www.jimmunol.org/ by guest on September 28, 2021 Inducing is online at: average * The Journal of Immunology published online 16 August 2013 from submission to initial decision 4 weeks from acceptance to publication http://www.jimmunol.org/content/early/2013/08/16/jimmun ol.1300376 A Synthetic TLR3 Ligand Mitigates Profibrotic Fibroblast Responses by Autocrine IFN Signaling Feng Fang, Kohtaro Ooka, Xiaoyong Sun, Ruchi Shah, Swati Bhattacharyya, Jun Wei and John Varga J Immunol Submit online. Every submission reviewed by practicing scientists ? is published twice each month by http://jimmunol.org/subscription Submit copyright permission requests at: http://www.aai.org/About/Publications/JI/copyright.html Receive free email-alerts when new articles cite this article. Sign up at: http://jimmunol.org/alerts http://www.jimmunol.org/content/suppl/2013/08/20/jimmunol.130037 6.DC1 Information about subscribing to The JI No Triage! Fast Publication! Rapid Reviews! 30 days* Why • • • Material Permissions Email Alerts Subscription Supplementary The Journal of Immunology The American Association of Immunologists, Inc., 1451 Rockville Pike, Suite 650, Rockville, MD 20852 Copyright © 2013 by The American Association of Immunologists, Inc. All rights reserved. Print ISSN: 0022-1767 Online ISSN: 1550-6606. This information is current as of September 28, 2021. Published August 16, 2013, doi:10.4049/jimmunol.1300376 The Journal of Immunology A Synthetic TLR3 Ligand Mitigates Profibrotic Fibroblast Responses by Inducing Autocrine IFN Signaling Feng Fang,* Kohtaro Ooka,* Xiaoyong Sun,† Ruchi Shah,* Swati Bhattacharyya,* Jun Wei,* and John Varga* Activation of TLR3 by exogenous microbial ligands or endogenous injury-associated ligands leads to production of type I IFN. -

Proteomic Landscape of the Human Choroid–Retinal Pigment Epithelial Complex
Supplementary Online Content Skeie JM, Mahajan VB. Proteomic landscape of the human choroid–retinal pigment epithelial complex. JAMA Ophthalmol. Published online July 24, 2014. doi:10.1001/jamaophthalmol.2014.2065. eFigure 1. Choroid–retinal pigment epithelial (RPE) proteomic analysis pipeline. eFigure 2. Gene ontology (GO) distributions and pathway analysis of human choroid– retinal pigment epithelial (RPE) protein show tissue similarity. eMethods. Tissue collection, mass spectrometry, and analysis. eTable 1. Complete table of proteins identified in the human choroid‐RPE using LC‐ MS/MS. eTable 2. Top 50 signaling pathways in the human choroid‐RPE using MetaCore. eTable 3. Top 50 differentially expressed signaling pathways in the human choroid‐RPE using MetaCore. eTable 4. Differentially expressed proteins in the fovea, macula, and periphery of the human choroid‐RPE. eTable 5. Differentially expressed transcription proteins were identified in foveal, macular, and peripheral choroid‐RPE (p<0.05). eTable 6. Complement proteins identified in the human choroid‐RPE. eTable 7. Proteins associated with age related macular degeneration (AMD). This supplementary material has been provided by the authors to give readers additional information about their work. © 2014 American Medical Association. All rights reserved. 1 Downloaded From: https://jamanetwork.com/ on 09/25/2021 eFigure 1. Choroid–retinal pigment epithelial (RPE) proteomic analysis pipeline. A. The human choroid‐RPE was dissected into fovea, macula, and periphery samples. B. Fractions of proteins were isolated and digested. C. The peptide fragments were analyzed using multi‐dimensional LC‐MS/MS. D. X!Hunter, X!!Tandem, and OMSSA were used for peptide fragment identification. E. Proteins were further analyzed using bioinformatics. -

Use of Chronic Lymphocytic Leukemia Research
USE OF CHRONIC LYMPHOCYTIC LEUKEMIA RESEARCH CONSORTIUM DATA REPOSITORY AND GENE EXPRESSION OMNIBUS TO GENERATE AND TEST HYPOTHESES FOR BIOMARKER IDENTIFICATION AND DEVELOPMENT THESIS Presented in Partial Fulfillment of the Requirements for the Degree Master of Science in the Graduate School of The Ohio State University By Kristin Chelsea Keen, B.A. ***** The Ohio State University 2009 Thesis Committee: Approved by Professor Kun Huang, adviser Professor Philip Payne ______________________________ Adviser Pathology Graduate Program ABSTRACT Chronic lymphocytic leukemia (CLL) is the most common adult leukemia in the United States. There is no known cure for CLL. While biomarkers have been found to correlate with disease progression, such as CD38, IGHV, and ZAP-70, there is a need for further validation of these biomarkers as well as new biomarker discovery. In this study, publicly available gene expression data from NCBI’s Gene Expression Omnibus was used to identify genes with expression correlated to ZAP-70 and CD38 mRNA expression patterns. Also utilized were vast amounts of data from the Chronic Lymphocytic Leukemia Research Consortium (CRC) to search for novel correlations between clinical markers and disease progression and treatment outcome. We found several hundred genes with expression patterns correlated to ZAP-70. We also found several clinical, genetic, biologic, and immunologic CRC data fields which were correlated significantly and at weakly to strongly associated. ii Dedicated to my family iii ACKNOWLEDGEMENTS I thank my adviser, Kun Huang, for his positive support and trust in me as a student and a researcher. I thank my committee member, Philip Payne, for his tangible enthusiasm and thoughtful advice. -

Biomedical Informatics
BIOMEDICAL INFORMATICS Abstract GENE LIST AUTOMATICALLY DERIVED FOR YOU (GLAD4U): DERIVING AND PRIORITIZING GENE LISTS FROM PUBMED LITERATURE JEROME JOURQUIN Thesis under the direction of Professor Bing Zhang Answering questions such as ―Which genes are related to breast cancer?‖ usually requires retrieving relevant publications through the PubMed search engine, reading these publications, and manually creating gene lists. This process is both time-consuming and prone to errors. We report GLAD4U (Gene List Automatically Derived For You), a novel, free web-based gene retrieval and prioritization tool. The quality of gene lists created by GLAD4U for three Gene Ontology terms and three disease terms was assessed using ―gold standard‖ lists curated in public databases. We also compared the performance of GLAD4U with that of another gene prioritization software, EBIMed. GLAD4U has a high overall recall. Although precision is generally low, its prioritization methods successfully rank truly relevant genes at the top of generated lists to facilitate efficient browsing. GLAD4U is simple to use, and its interface can be found at: http://bioinfo.vanderbilt.edu/glad4u. Approved ___________________________________________ Date _____________ GENE LIST AUTOMATICALLY DERIVED FOR YOU (GLAD4U): DERIVING AND PRIORITIZING GENE LISTS FROM PUBMED LITERATURE By Jérôme Jourquin Thesis Submitted to the Faculty of the Graduate School of Vanderbilt University in partial fulfillment of the requirements for the degree of MASTER OF SCIENCE in Biomedical Informatics May, 2010 Nashville, Tennessee Approved: Professor Bing Zhang Professor Hua Xu Professor Daniel R. Masys ACKNOWLEDGEMENTS I would like to express profound gratitude to my advisor, Dr. Bing Zhang, for his invaluable support, supervision and suggestions throughout this research work.