Quantitatively Modeling Factors That Influence the Therapeutic Doses Of
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
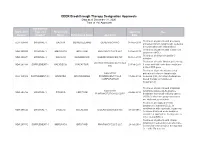
CDER Breakthrough Therapy Designation Approvals Data As of December 31, 2020 Total of 190 Approvals
CDER Breakthrough Therapy Designation Approvals Data as of December 31, 2020 Total of 190 Approvals Submission Application Type and Proprietary Approval Use Number Number Name Established Name Applicant Date Treatment of patients with previously BLA 125486 ORIGINAL-1 GAZYVA OBINUTUZUMAB GENENTECH INC 01-Nov-2013 untreated chronic lymphocytic leukemia in combination with chlorambucil Treatment of patients with mantle cell NDA 205552 ORIGINAL-1 IMBRUVICA IBRUTINIB PHARMACYCLICS LLC 13-Nov-2013 lymphoma (MCL) Treatment of chronic hepatitis C NDA 204671 ORIGINAL-1 SOVALDI SOFOSBUVIR GILEAD SCIENCES INC 06-Dec-2013 infection Treatment of cystic fibrosis patients age VERTEX PHARMACEUTICALS NDA 203188 SUPPLEMENT-4 KALYDECO IVACAFTOR 21-Feb-2014 6 years and older who have mutations INC in the CFTR gene Treatment of previously untreated NOVARTIS patients with chronic lymphocytic BLA 125326 SUPPLEMENT-60 ARZERRA OFATUMUMAB PHARMACEUTICALS 17-Apr-2014 leukemia (CLL) for whom fludarabine- CORPORATION based therapy is considered inappropriate Treatment of patients with anaplastic NOVARTIS lymphoma kinase (ALK)-positive NDA 205755 ORIGINAL-1 ZYKADIA CERITINIB 29-Apr-2014 PHARMACEUTICALS CORP metastatic non-small cell lung cancer (NSCLC) who have progressed on or are intolerant to crizotinib Treatment of relapsed chronic lymphocytic leukemia (CLL), in combination with rituximab, in patients NDA 206545 ORIGINAL-1 ZYDELIG IDELALISIB GILEAD SCIENCES INC 23-Jul-2014 for whom rituximab alone would be considered appropriate therapy due to other co-morbidities -

Pub 100-04 Medicare Claims Processing Centers for Medicare & Medicaid Services (CMS) Transmittal 4410 Date: October 4, 2019 Change Request 11457
Department of Health & CMS Manual System Human Services (DHHS) Pub 100-04 Medicare Claims Processing Centers for Medicare & Medicaid Services (CMS) Transmittal 4410 Date: October 4, 2019 Change Request 11457 Transmittal 4389, dated September 6, 2019, is being rescinded and replaced by Transmittal 4410, dated, October 4, 2019 to correct table 1 attachment A to reinstate C9043 rather than delete it effective October 1, 2019, revise corresponding footnote 1, and delete footnote 2. Filenames have been corrected for BR11457.1 and BR11457.2. All other information remains the same. SUBJECT: October 2019 Update of the Ambulatory Surgical Center (ASC) Payment System I. SUMMARY OF CHANGES: This Recurring Update Notification (RUN) provides instructions for the quarterly update to the Ambulatory Surgical Center (ASC) Payment System. EFFECTIVE DATE: October 1, 2019 *Unless otherwise specified, the effective date is the date of service. IMPLEMENTATION DATE: October 7, 2019 Disclaimer for manual changes only: The revision date and transmittal number apply only to red italicized material. Any other material was previously published and remains unchanged. However, if this revision contains a table of contents, you will receive the new/revised information only, and not the entire table of contents. II. CHANGES IN MANUAL INSTRUCTIONS: (N/A if manual is not updated) R=REVISED, N=NEW, D=DELETED-Only One Per Row. R/N/D CHAPTER / SECTION / SUBSECTION / TITLE N/A N/A III. FUNDING: For Medicare Administrative Contractors (MACs): The Medicare Administrative Contractor is hereby advised that this constitutes technical direction as defined in your contract. CMS does not construe this as a change to the MAC Statement of Work. -

Antibodies to Watch in 2021 Hélène Kaplona and Janice M
MABS 2021, VOL. 13, NO. 1, e1860476 (34 pages) https://doi.org/10.1080/19420862.2020.1860476 PERSPECTIVE Antibodies to watch in 2021 Hélène Kaplona and Janice M. Reichert b aInstitut De Recherches Internationales Servier, Translational Medicine Department, Suresnes, France; bThe Antibody Society, Inc., Framingham, MA, USA ABSTRACT ARTICLE HISTORY In this 12th annual installment of the Antibodies to Watch article series, we discuss key events in antibody Received 1 December 2020 therapeutics development that occurred in 2020 and forecast events that might occur in 2021. The Accepted 1 December 2020 coronavirus disease 2019 (COVID-19) pandemic posed an array of challenges and opportunities to the KEYWORDS healthcare system in 2020, and it will continue to do so in 2021. Remarkably, by late November 2020, two Antibody therapeutics; anti-SARS-CoV antibody products, bamlanivimab and the casirivimab and imdevimab cocktail, were cancer; COVID-19; Food and authorized for emergency use by the US Food and Drug Administration (FDA) and the repurposed Drug Administration; antibodies levilimab and itolizumab had been registered for emergency use as treatments for COVID-19 European Medicines Agency; in Russia and India, respectively. Despite the pandemic, 10 antibody therapeutics had been granted the immune-mediated disorders; first approval in the US or EU in 2020, as of November, and 2 more (tanezumab and margetuximab) may Sars-CoV-2 be granted approvals in December 2020.* In addition, prolgolimab and olokizumab had been granted first approvals in Russia and cetuximab saratolacan sodium was first approved in Japan. The number of approvals in 2021 may set a record, as marketing applications for 16 investigational antibody therapeutics are already undergoing regulatory review by either the FDA or the European Medicines Agency. -

Magro Virus Research 2020
Virus Research 286 (2020) 198070 Contents lists available at ScienceDirect Virus Research journal homepage: www.elsevier.com/locate/virusres Review COVID-19: Review on latest available drugs and therapies against SARS- T CoV-2. Coagulation and inflammation cross-talking Giuseppe Magro Department of Medical, Surgical Sciences, University "Magna Græcia" of Catanzaro, Italy ARTICLE INFO ABSTRACT Keywords: SARS-CoV-2 is the agent responsible for COVID-19. The infection can be dived into three phases: mild infection, SARS-CoV-2 the pulmonary phase and the inflammatory phase. Treatment options for the pulmonary phase include: COVID-19 Hydroxychloroquine, Remdesivir, Lopinavir/Ritonavir. The inflammatory phase includes therapeutic options Therapies like Tocilizumab, Anakinra, Baricitinib, Eculizumab, Emapalumab and Heparin. Human clinical trials are Drugs starting to show some results, in some cases like that of Remdesivir and corticosteroids these are controversial. SARS-CoV-1 Coagulopathy is a common complication in severe cases, inflammation and coagulation are intertwined and Coronavirus Treatment cross-talking between these two responses is known to happen. A possible amplification of this cross-talking is Tocilizumab suggested to be implicated in the severe cases that show both a cytokine storm and coagulopathy. Remdesivir Emapalumab Eculizumab Ruxolitinib Baricitinib Dexamethasone Coagulation Inflammation 1. Introduction diarrhea (Wang et al., 2020a), confusion, sore throat, rhinorrhea, chest pain, nausea and vomiting (Chen et al., 2020a). Up to 50 % of patients Coronaviruses are enveloped, positive-sense, single stranded RNA develop shortness of breath. Acute respiratory distress syndrome viruses that are distributed broadly among humans which cause re- (ARDS) is not an uncommon complication when disease cannot be spiratory, enteric, hepatic, and neurologic diseases (Weiss and controlled (She et al., 2020). -

Swiss Biotech Report – "Agility, Leadership and Innovation in the Time of COVID-19” – Is Indeed Appropriate
SwissSwiss BiotechBiotech ReportReport 2021 2019Agility, leadership and innovation in the time of COVID-19 Schweizerische Eidgenossenschaft Confédération suisse Confederazione Svizzera Confederaziun svizra Swiss Confederation Innosuisse – Swiss Innovation Agency “The biopharma industry’s response to COVID-19 has truly been a global one – with Swiss companies and academic institutions at the forefront. Swiss-based biopharma players are contributing across the spectrum of patient needs.” George Scangos | Vir Biotechnology, Inc. “2020 was the best year ever for Swiss biotech in relation to financing activities, with a total of approximately CHF 3.4B raised” Jürg Zürcher | GSA Biotechnology Leader, EY “Compared to 2019, trading volumes rose sharply by 177% and the total free float market capitalization of all SIX-listed biotech companies was up 30% on 2020.” Fabian Gerber | SIX Agility, leadership and innovation in the time of COVID-19 “Technologies developed by scientists in Switzerland have been crucial in the fight against COVID-19, including cloning the virus, cryo electron microscopy to visualize the spike proteins, and the use of radioactive molecules that bind to receptor proteins to investigate how the virus enters the host.” Florian Fisch | Swiss National Science Foundation “On May 1 2020, Moderna and Lonza announced a strategic collaboration to enable manufacturing of up to 1 billion doses of the vaccine per year. Lonza acquired equipment and built the production lines in just 8 months.” Jan Lucht | scienceindustries “Industrial -

CDER Therapeutic Biologic Products List
CDER Therapeutic Biologic Products This list is intended to include all the Center for Drug Evaluation and Research (CDER) user fee billable therapeutic biological products and potencies approved under Section 351 of the Public Health Service Act. The Orange Book includes a section entitled "Drug Products with Approval under Section 505 of the Act Administered by CBER." Included on that list are several products that have been transferred to CDER which would be considered billable also. Program fees are assessed for each potency in which the approved (non-revoked, non-suspended) product is manufactured in final dosage form. When evaluating the specific strength or potency of a drug in final dosage form for purposes of assessing program fees for liquid parenteral biological products, CDER intends to take into consideration both the total amount of drug substance in mass or units of activity in a product and the concentration of drug substance (mass or units of activity per unit volume of product). Biologic products considered to have a different strength or potency in a final dosage form will be given separate entries in the Biologics List and assessed separate program fees. An auto-injector that has the same strength or potency as a prefilled syringe or vial will generally be assessed a separate prescription drug program fee. In certain circumstances, products which have been discontinued from marketing but are still licensed are not assessed program fees. Those products are identified on the CDER Discontinued Biologic Product List section. The potency information contained in this list is based on information in our database. -

Quartz Medicare Advantage 2021 Part B Prior Authorization Criteria
Quartz Medicare Advantage (HMO) Part B Medication (Medical Benefit) Prior Authorization Criteria These medication prior authorization criteria apply to Quartz Medicare Advantage members for medications when given by a health care provider (medical benefit) and billed under Medicare Part B. GH00432 (1020) Y0092_20 117_C Afamelanotide (Scenesse) Prior Authorization Criteria Drug Name Drug Status Quantity Limits Approval Limits Afamelanotide (Scenesse) Medical benefit- None- implant 12 months Restricted every 2 months CRITERIA FOR COVERAGE: • Diagnosis of Erythropoietic Protoporphyria (EPP), AND • Age ≥18 years, AND • History of phototoxic reactions due to free light exposure CRITERIA FOR CONTINUATION OF THERAPY: (for renewal or new members. This criteria will be applied if the requested medication has been used in the previous 365 days) • Initial criteria met and clinical documentation from the previous 12 months demonstrating objective improvements in pain control related to light exposure • Continuation of therapy/coverage criteria will not be applied to persons who were not previously approved for coverage whose therapy was initiated using a manufacturer-sponsored free drug program, provider samples, and/or vouchers. (7/1/2021) Quartz Medicare Advantage 1 Agalsidase Beta (Fabrazyme) Prior Authorization Criteria Drug Name Drug Status Quantity Limits Approval Limits Agalsidase Beta Medical Benefit- 1mg/kg IV infusion None (Fabrazyme) Restricted every two weeks CRITERIA FOR COVERAGE: • Diagnosis of Fabry’s Disease, AND • Prescribed by or -

Soluble Ligands As Drug Targets
REVIEWS Soluble ligands as drug targets Misty M. Attwood 1, Jörgen Jonsson1, Mathias Rask- Andersen 2 and Helgi B. Schiöth 1,3 ✉ Abstract | Historically, the main classes of drug targets have been receptors, enzymes, ion channels and transporters. However, owing largely to the rise of antibody- based therapies in the past two decades, soluble protein ligands such as inflammatory cytokines have become an increasingly important class of drug targets. In this Review, we analyse drugs targeting ligands that have reached clinical development at some point since 1992. We identify 291 drugs that target 99 unique ligands, and we discuss trends in the characteristics of the ligands, drugs and indications for which they have been tested. In the last 5 years, the number of ligand-targeting drugs approved by the FDA has doubled to 34, while the number of clinically validated ligand targets has doubled to 22. Cytokines and growth factors are the predominant types of targeted ligands (70%), and inflammation and autoimmune disorders, cancer and ophthalmological diseases are the top therapeutic areas for both approved agents and agents in clinical studies, reflecting the central role of cytokine and/or growth factor pathways in such diseases. Drug targets In the twentieth century, drug discovery largely involved far more challenging to achieve with small- molecule Pharmacological targets, such the identification of small molecules that exert their drugs. Protein ligands have been successfully targeted as proteins, that mediate the therapeutic effects by interacting with the binding sites by many drugs since the first FDA approval of the desired therapeutic effect of of endogenous small- molecule ligands such as neuro- ligand- targeting agents etanercept and infliximab in a drug. -

Minutes of the CHMP Meeting 9-12 November 2020
05 January 2021 EMA/CHMP/708314/2020 Human Medicines Division Committee for medicinal products for human use (CHMP) Minutes for the meeting on 09-12 November 2020 Chair: Harald Enzmann – Vice-Chair: Bruno Sepodes Disclaimers Some of the information contained in these minutes is considered commercially confidential or sensitive and therefore not disclosed. With regard to intended therapeutic indications or procedure scopes listed against products, it must be noted that these may not reflect the full wording proposed by applicants and may also vary during the course of the review. Additional details on some of these procedures will be published in the CHMP meeting highlights once the procedures are finalised and start of referrals will also be available. Of note, these minutes are a working document primarily designed for CHMP members and the work the Committee undertakes. Note on access to documents Some documents mentioned in the minutes cannot be released at present following a request for access to documents within the framework of Regulation (EC) No 1049/2001 as they are subject to on- going procedures for which a final decision has not yet been adopted. They will become public when adopted or considered public according to the principles stated in the Agency policy on access to documents (EMA/127362/2006). Official address Domenico Scarlattilaan 6 ● 1083 HS Amsterdam ● The Netherlands Address for visits and deliveries Refer to www.ema.europa.eu/how-to-find-us Send us a question Go to www.ema.europa.eu/contact Telephone +31 (0)88 781 6000 An agency of the European Union © European Medicines Agency, 2020. -

Overview of Planned Or Ongoing Studies of Drugs for the Treatment of COVID-19
Version of 16.06.2020 Overview of planned or ongoing studies of drugs for the treatment of COVID-19 Table of contents Antiviral drugs ............................................................................................................................................................. 4 Remdesivir ......................................................................................................................................................... 4 Lopinavir + Ritonavir (Kaletra) ........................................................................................................................... 7 Favipiravir (Avigan) .......................................................................................................................................... 14 Darunavir + cobicistat or ritonavir ................................................................................................................... 18 Umifenovir (Arbidol) ........................................................................................................................................ 19 Other antiviral drugs ........................................................................................................................................ 20 Antineoplastic and immunomodulating agents ....................................................................................................... 24 Convalescent Plasma ........................................................................................................................................... -

T Cell-Epstein-Barr Virus–Associated Hemophagocytic Lymphohistiocytosis (T Cell-EBV-HLH) Is Prevalent in East Asia and Has Poor Prognosis
Journal of Clinical Immunology (2021) 41:1582–1596 https://doi.org/10.1007/s10875-021-01073-9 ORIGINAL ARTICLE T Cell‑Epstein‑Barr Virus–Associated Hemophagocytic Lymphohistiocytosis (HLH) Occurs in Non‑Asians and Is Associated with a T Cell Activation State that Is Comparable to Primary HLH Oded Shamriz1,2 · Deepak Kumar1,2 · Jenny Shim1,2 · Michael Briones1,2 · Maa‑Ohui Quarmyne1,3 · Satheesh Chonat1,2 · Laura Lucas4 · Holly Edington1,2 · Michael H. White1,2 · Advay Mahajan1,2 · Sunita Park5 · Shanmuganathan Chandrakasan1 Received: 31 March 2021 / Accepted: 20 May 2021 / Published online: 26 June 2021 © The Author(s), under exclusive licence to Springer Science+Business Media, LLC, part of Springer Nature 2021 Abstract Purpose T cell-Epstein-Barr virus–associated hemophagocytic lymphohistiocytosis (T cell-EBV-HLH) is prevalent in East Asia and has poor prognosis. Understanding of this disease is limited, and literature regarding prevalence in North America is scarce. Herein, we summarize our experience. Methods A retrospective analysis of T cell-EBV-HLH patients admitted to Children’s Healthcare of Atlanta (GA, USA) from 2010 to 2020 was conducted. Additional immune studies were completed in a subset of patients. Results We report 15 patients (10 months–19 years of age) diagnosed with T cell-EBV-HLH. Nine patients were Hispanic, and the majority did not have primary HLH (p-HLH) gene defects. Soluble interleukin-2 receptor levels in T cell-EBV-HLH were signifcantly higher than other forms of secondary-HLH but comparable to p-HLH, and it correlated with disease severity at presentation. Natural killer cell function was decreased in most patients despite a negative workup for p-HLH. -

Picture As Pdfoctober 2020: Reminder Specific Medications
OCTOBER 2020: REMINDER SPECIFIC MEDICATIONS REQUIRE PRIOR AUTHORIZATION As a part of our continuous efforts to improve the quality of care for our members, Highmark Health Options wanted to send a reminder about the prior authorization process for the following medications. Failure to obtain authorization will result in a claim denial. The prior authorization process applies to all Highmark Health Options Members. Medical necessity criteria for each of the medications listed below are outlined in the specific medication policies available online. To access Highmark Health Options’ medical policies, please paste the following link in your internet browser: https://www.highmarkhealthoptions.com/providers/medication-information.html. PROCEDURE CODES REQUIRING AUTHORIZATION DRUG NAME CODE DESCRIPTION HCPCS DE Medicaid Injection, abatacept, 10 mg (code may be used for Medicare when drug administered under J0129 11/4/2019 the direct supervision of a physician, not for abatacept (Orencia) use when drug is self administered aflibercept (Eylea) Injection, aflibercept, 1 mg J0178 10/1/2018 agalsidase beta (Fabrazyme) Injection, agalsidase beta, 1 mg J0180 6/3/2019 alemtuzumab (Lemtrada) Injection, alemtuzumab, 1 mg J0202 11/4/2019 alglucosidase alfa (Lumizyme) Injection, alglucosidase alfa, (lumizyme), 10 mg J0221 10/1/2018 aprepitant (Cinvanti) Injection, aprepitant, 1 mg J0185 11/4/2019 asparaginase (Erwinaze) Injection, asparaginase (erwinaze), 1,000 iu J9019 11/4/2019 atezolizumab (Tecentriq) Injection, atezolizumab, 10 mg J9022 11/4/2019 Axicabtagene