Zoölogisch Museum
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Observaciones Ornitológicas En El Archipiélago De Cabo Verde
Revista Academia Canaria de las Ciencias, X (Numero 4), 41-64 (1998) OBSERVACIONES ORNITOLOGICAS EN EL ARCHIPIELAGO DE CABO VERDE, SEPTJEMBRE-OCTUBRE DE 1998 D. AVES NIDIFICANTES (*) R. Barone* & G. Delgado** *C/. Eduardo Zamacois, 13-3°A, E-38005 Santa Cruz de Tenerife, Islas Canarias **Museo de la Naturaleza y el Hombre (Ciencias Naturales). Apartado. 853, E-38003 Santa Cruz de Tenerife, islas Canarias ABSTRACT Recent data on the breeding birds of the islands of Sal and Boavista (Cape Verde Islands), obtained between 30 September - 14 October, 1998, are presented. The nesting of the Red-billed Tropicbird (Phaethon aethe reus) on Boavista is confirmed, and the first registers of the Barn Owl {Tyto alba ssp.) for this island are included here. On the other hand, we could detect species found only 2-3 times on Boavista, as the Black Kite (Milvus migrans) and the Blackcap (Sylvia atricapilla). In the case of this last species, our data indi- cate that it must be a breeding species on this island. At the same time, several aspects of the avifauna of both islands are analysed: faunistic, habitats, breeding phenology and impor- tant areas for conservation. Key words: Breeding birds, Sal, Boavista, Cape Verde Islands, new records, faunistic, habitats, breeding phenology, conservation. RESUMEN Se presentan datos recientes sobre la avifauna nidificante en las islas de Sal y Boavista (archipielago de Cabo Verde), obtenidos entre el 30 de septiembre y el 14 de octubre de 1998. Destaca la confirmacion de la nidificacion del Rabijunco Etereo {Phaethon aethereus) en Boavista, asi como los primeros registros de Lechuza Comun (Tyto alba ssp.) para esta isla. -

Systematic List of the Birds
59.82(66.53) Article III.-THE MARINE ORNITHOLOGY OF THE CAPE VERDE ISLANDS, WITH A LIST OF ALL THE BIRDS OF THE ARCHIPELAGO BY ROBERT CUSHMAN MURPHY CONTENTS PAGE. INTRODUCTION........................................................... 211 HISTORICAL SKETCH OF CAPE VERDE ORNITHOLOGY .............. .......... 212 THE VISITS OF CORREIA AND THE AUTHOR.................................. 213 THE GEOGRAPHIC ENVIRONMENT....................... 216 OCEANOGRAPHY AND CLIMATE............................................ 221 GEOGRAPHIC RELATIONSHIPS OF THE BIRDS ............................... 226 BIBLIOGRAPHY......................................................... 232 SYSTEMATIC LIST OF THE BIRDS............................................ 233 INTRODUCTION The present paper is an indirect result of a brief visit which the writer made to the Cape Verde Islands in September 1912. It is more immediately linked, however, with a collecting trip undertaken during part of the spring and summer of 1922 by Mr. Jos6 Gongalves Correia, of New Bedford, Mass., whose specimens, numbering some three hundred skins of twenty species, have all been received by The American Museum of Natural History. As Correia's predominant object was to acquire adequate series of water birds, the terrestrial species of the Cape Verdes are but scantily represented in his collection, which is supplemented, to a certain extent, by a small number of specimens obtained by the writer in 1912. Notwithstanding the relatively limited material available for study, it has been thought advisable to include within this paper a systematic list of all the birds known from the Cape Verde Islands. The latest faunal contribution is that of Salvadori (1899). Not only have addi- tions to the avifauna been made since that date, but also forms new to science, including several endemic races, have been described. More- over, numerous nomenclatural changes made necessary because of the researches of Hartert ('V6gel der palaarktischen Fauna,' 1910-22) and other workers increase the need for a new list. -

FEEDING ECOLOGY of the CAPE VERDEAN SHEARWATER (Calonectris Edwardsii) POPULATION of RASO ISLET, CAPE VERDE (P1-B-11)
2nd World Seabird Conference “Seabirds: Global Ocean Sentinels” 26-30 October 2015 Cape Town, South Africa FEEDING ECOLOGY OF THE CAPE VERDEAN SHEARWATER (Calonectris edwardsii) POPULATION OF RASO ISLET, CAPE VERDE (P1-B-11) Isabel Rodrigues1,3; Nuno Oliveira 2 Rui Freitas 3 Tommy Melo1 Pedro Geraldes 2 1Biosfera I, Cabo Verde, www.biosfera1.com; 2SPEA, Portugal, www.spea.pt, 3 Universidade de Cabo Verde, www.unicv.edu.cv Raso Islet with only 5.76 km2, is an area of great importance to Cape Verdean Shearwaters as we find one of the largest colonies of the species there. The great biological value of this islet is even more remarkable for hosting very large populations of other species, such as the Brown Booby, Red-billed Tropicalbird and even the endemic Raso Lark, among others. Along with Branco Islet, also included in the Nature Reserve, both populations constitute about 75% of the nesting population of the Cape Verde islands (Fig. 1). The Cape Verde Shearwater (Procellariiformes, Procellariidae) (Fig. 2) is an endemic species of Cape Verde and has recently been separated from Calonectris diomedea species, due to their morphological and genetic differences, and pelagic habits; feeding mostly on the open sea (Hazevoet 1995). METHODOLOGY RESULTS The samples were collected between 14 October and 12 November, in two In total, 80 regurgitations from juvenile Cape Verde Shearwaters were collected; consecutive years, 2012 and 2013. We randomly obtained 80 samples of juvenile including 50 individuals sampled in 2012, and 30 individuals in 2013. regurgitation. Each juvenile was sampled only once. During or after handling, Based on knowledge of local fish populations and according to the current juveniles tend to regurgitate stomach contents without the need to resort to the description, the identified prey of Cape Verde shearwaters belonged to 5 species induced regurgitation method. -

In the Cape Verde Islands
ZOOLOGIA CABOVERDIANA REVISTA DA SOCIEDADE CABOVERDIANA DE ZOOLOGIA VOLUME 5 | NÚMERO 1 Abril de 2014 ZOOLOGIA CABOVERDIANA REVISTA DA SOCIEDADE CABOVERDIANA DE ZOOLOGIA Zoologia Caboverdiana is a peer-reviewed open-access journal that publishes original research articles as well as review articles and short notes in all areas of zoology and paleontology of the Cape Verde Islands. Articles may be written in English (with Portuguese summary) or Portuguese (with English summary). Zoologia Caboverdiana is published biannually, with issues in spring and autumn. For further information, contact the Editor. Instructions for authors can be downloaded at www.scvz.org Zoologia Caboverdiana é uma revista científica com arbitragem científica (peer-review) e de acesso livre. Nela são publicados artigos de investigação original, artigos de síntese e notas breves sobre zoologia e paleontologia das Ilhas de Cabo Verde. Os artigos podem ser submetidos em inglês (com um resumo em português) ou em português (com um resumo em inglês). Zoologia Caboverdiana tem periodicidade bianual, com edições na primavera e no outono. Para mais informações, deve contactar o Editor. Normas para os autores podem ser obtidas em www.scvz.org Chief Editor | Editor principal Dr Cornelis J. Hazevoet (Instituto de Investigação Científica Tropical, Portugal); [email protected] Editorial Board | Conselho editorial Dr Joana Alves (Instituto Nacional de Saúde Pública, Praia, Cape Verde) Prof. Dr G.J. Boekschoten (Vrije Universiteit Amsterdam, The Netherlands) Dr Eduardo Ferreira (Universidade de Aveiro, Portugal) Rui M. Freitas (Universidade de Cabo Verde, Mindelo, Cape Verde) Dr Javier Juste (Estación Biológica de Doñana, Spain) Evandro Lopes (Universidade de Cabo Verde, Mindelo, Cape Verde) Dr Adolfo Marco (Estación Biológica de Doñana, Spain) Prof. -

Recent Bird Records from Fogo, Cape Verde Islands Rubén Baronea and Jens Heringb
Recent bird records from Fogo, Cape Verde Islands Rubén Baronea and Jens Heringb Observations récentes de Fogo, Îles du Cap-Vert. Des données sont présentées concernant 12 espèces d’oiseaux observées à Fogo, Îles du Cap-Vert, parmi lesquelles deux premières mentions pour l’île (Chevalier gambette Tringa totanus et Hirondelle de fenêtre Delichon urbicum), les premières données de nidification du Martinet du Cap-Vert Apus alexandri et les premières observations fiables du Phaéton à bec rouge Phaethon aethereus indiquant la nidification probable de celui-ci. Des informations sont également présentées sur d’autres taxons mal connus à Fogo, tels que certaines espèces pélagiques et l’Effraie des clochers Tyto alba detorta. Summary. We present data on 12 bird species observed on Fogo, Cape Verde Islands, among them two first records for the island (Common Redshank Tringa totanus and Common House Martin Delichon urbicum), the first breeding records of Cape Verde Swift Apus alexandri and the first reliable observations of Red-billed Tropicbird Phaethon aethereus indicating probable breeding. Information on other taxa poorly known on Fogo, such as some pelagic seabirds and Barn Owl Tyto alba detorta, is also given. ogo, one of the Cape Verde Islands, is situated reported previously, and some others for which F in the leeward group (‘Ilhas do Sotavento’), there are only a limited number of observations. c.724 km from the African continent. With Local information on breeding birds was mainly a surface area of 478 km2, the highest peak collected by RB. Dates of our visits are as follows: (Pico Novo) reaches 2,829 m (Michell-Thomé 18–21 October 2004 (JH & H. -
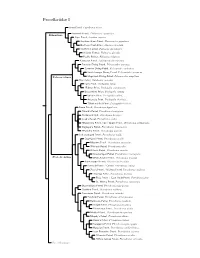
Procellariidae Species Tree
Procellariidae I Snow Petrel, Pagodroma nivea Antarctic Petrel, Thalassoica antarctica Fulmarinae Cape Petrel, Daption capense Southern Giant-Petrel, Macronectes giganteus Northern Giant-Petrel, Macronectes halli Southern Fulmar, Fulmarus glacialoides Atlantic Fulmar, Fulmarus glacialis Pacific Fulmar, Fulmarus rodgersii Kerguelen Petrel, Aphrodroma brevirostris Peruvian Diving-Petrel, Pelecanoides garnotii Common Diving-Petrel, Pelecanoides urinatrix South Georgia Diving-Petrel, Pelecanoides georgicus Pelecanoidinae Magellanic Diving-Petrel, Pelecanoides magellani Blue Petrel, Halobaena caerulea Fairy Prion, Pachyptila turtur ?Fulmar Prion, Pachyptila crassirostris Broad-billed Prion, Pachyptila vittata Salvin’s Prion, Pachyptila salvini Antarctic Prion, Pachyptila desolata ?Slender-billed Prion, Pachyptila belcheri Bonin Petrel, Pterodroma hypoleuca ?Gould’s Petrel, Pterodroma leucoptera ?Collared Petrel, Pterodroma brevipes Cook’s Petrel, Pterodroma cookii ?Masatierra Petrel / De Filippi’s Petrel, Pterodroma defilippiana Stejneger’s Petrel, Pterodroma longirostris ?Pycroft’s Petrel, Pterodroma pycrofti Soft-plumaged Petrel, Pterodroma mollis Gray-faced Petrel, Pterodroma gouldi Magenta Petrel, Pterodroma magentae ?Phoenix Petrel, Pterodroma alba Atlantic Petrel, Pterodroma incerta Great-winged Petrel, Pterodroma macroptera Pterodrominae White-headed Petrel, Pterodroma lessonii Black-capped Petrel, Pterodroma hasitata Bermuda Petrel / Cahow, Pterodroma cahow Zino’s Petrel / Madeira Petrel, Pterodroma madeira Desertas Petrel, Pterodroma -

Shearwatersrefs V1.10.Pdf
Introduction I have endeavoured to keep typos, errors, omissions etc in this list to a minimum, however when you find more I would be grateful if you could mail the details during 2018 & 2019 to: [email protected]. Please note that this and other Reference Lists I have compiled are not exhaustive and are best employed in conjunction with other sources. Grateful thanks to Ashley Fisher (www.scillypelagics.com) for the cover images. All images © the photographer. Joe Hobbs Index The general order of species follows the International Ornithologists' Union World Bird List (Gill, F. & Donsker, D. (eds.) 2017. IOC World Bird List. Available from: http://www.worldbirdnames.org/ [version 7.3 accessed August 2017]). Version Version 1.10 (January 2018). Cover Main image: Great Shearwater. At sea 3’ SW of the Bishop Rock, Isles of Scilly. 8th August 2009. Picture by Ashley Fisher. Vignette: Sooty Shearwater. At sea off the Isles of Scilly. 14th August 2009. Picture by Ashley Fisher. Species Page No. Audubon's Shearwater [Puffinus lherminieri] 34 Balearic Shearwater [Puffinus mauretanicus] 28 Bannerman's Shearwater [Puffinus bannermani] 37 Barolo Shearwater [Puffinus baroli] 38 Black-vented Shearwater [Puffinus opisthomelas] 30 Boyd's Shearwater [Puffinus boydi] 38 Bryan's Shearwater [Puffinus bryani] 29 Buller's Shearwater [Ardenna bulleri] 15 Cape Verde Shearwater [Calonectris edwardsii] 12 Christmas Island Shearwater [Puffinus nativitatis] 23 Cory's Shearwater [Calonectris borealis] 9 Flesh-footed Shearwater [Ardenna carneipes] 21 Fluttering -

Threats to Seabirds: a Global Assessment 2 3 4 Authors: Maria P
1 Threats to seabirds: a global assessment 2 3 4 Authors: Maria P. Dias1*, Rob Martin1, Elizabeth J. Pearmain1, Ian J. Burfield1, Cleo Small2, Richard A. 5 Phillips3, Oliver Yates4, Ben Lascelles1, Pablo Garcia Borboroglu5, John P. Croxall1 6 7 8 Affiliations: 9 1 - BirdLife International. The David Attenborough Building, Pembroke Street Cambridge CB2 3QZ UK 10 2 - BirdLife International Marine Programme, RSPB, The Lodge, Sandy, SG19 2DL 11 3 – British Antarctic Survey. Natural Environment Research Council, High Cross, Madingley Road, 12 Cambridge CB3 0ET, UK 13 4 – Centre for the Environment, Fishery and Aquaculture Science, Pakefield Road, Lowestoft, NR33, UK 14 5 - Global Penguin Society, University of Washington and CONICET Argentina. Puerto Madryn U9120, 15 Chubut, Argentina 16 * Corresponding author: Maria Dias, [email protected]. BirdLife International. The David 17 Attenborough Building, Pembroke Street Cambridge CB2 3QZ UK. Phone: +44 (0)1223 747540 18 19 20 Acknowledgements 21 We are very grateful to Bartek Arendarczyk, Sophie Bennett, Ricky Hibble, Eleanor Miller and Amy 22 Palmer-Newton for assisting with the bibliographic review. We thank Rachael Alderman, Pep Arcos, 23 Jonathon Barrington, Igor Debski, Peter Hodum, Gustavo Jimenez, Jeff Mangel, Ken Morgan, Paul Sagar, 24 Peter Ryan, and other members of the ACAP PaCSWG, and the members of IUCN SSC Penguin Specialist 25 Group (Alejandro Simeone, Andre Chiaradia, Barbara Wienecke, Charles-André Bost, Lauren Waller, Phil 26 Trathan, Philip Seddon, Susie Ellis, Tom Schneider and Dee Boersma) for reviewing threats to selected 27 species. We thank also Andy Symes, Rocio Moreno, Stuart Butchart, Paul Donald, Rory Crawford, 28 Tammy Davies, Ana Carneiro and Tris Allinson for fruitful discussions and helpful comments on earlier 29 versions of the manuscript. -

Trip Report Cape Verde
Cape Verde March 17 th – 24 th 2018 Cape Verde Swamp Warbler Red-billed Tropicbird Raso Lark Islands (and sites) visited: Santiago Praia cliffs, Praia plains, Barragem de Poilão, Pedra Badejo lagoons, Barragem de Figuera Gorda, Jardim Botanico Boavista Sal Rei harbor, Rabil lagoon, Ponta da Varandinha, Zone Humide de Lecacão, Curral Velho, centre of isle São Nicolau Ponta do Barril, Taraffal Raso General Information Getting there: Cape Verde lies approx. 600 km west of Senegal and consists of 9 inhabited plus a number of un- inhabited islands. Thus flying there is the most common way to reach Cape Verde. Most visitors will be landing on Sal or Boavista as these are the most touristic islands and several airlines go there. Santiago and Sao Vincente also have international airports. TAP heads there via Lisbon. A visa is required for EU-citizens which can be obtained in the arrival airport for 27€. This proce- dure requires some time, depending on the amount of passengers in the respective plane. Visas can also be requested in the Cape Verde embassies in most countries. This was 45€ for Germany. Getting around: Between several islands: If you want to visit more than one island the local airline Binter offers flights for reasonable prices. They are said to not always stick to the schedule but were very reli- able in my case. Some islands can be reached by ferry but be aware that during the windy season in winter and spring the sea can be very rough and many people get seasick. On the islands: This proves to be one of the expensive parts. -

Study and Conservation of Breeding Seabirds in Rabo-De-Junco Islet Sal Island, Cape Verde
Associação Projeto Biodiversidade (Project Biodiversity) Albert Taxonera & Marcos Hernández Study and Conservation of breeding Seabirds in Rabo-de-junco Islet Sal Island, Cape Verde Project Biodiveristy Project Biodiversity is a Cabo Verdean organization committed to conserving and restoring the island’s unique ecosystems. Based on the island of Sal, the project implements community- based initiatives that promote conservation and better understanding of the island’s natural resources while increasing economic opportunities for the growing local community. Visit www.projectbiodiversity.org to know more about our initiatives. Seabird species of Sal Cabo Verde is a volcanic archipelago of 10 island and several islets included in the Macaronesia region, and located over 570 km off the coast of West Africa. The Island of Sal is one of the nine inhabited islands and located in the northeast corner of the archipelago (Figure 1). It is fairly flat with an area of 216 km2 and its highest point culminates at 408 meters. The southern part of the island, where sandy beaches extend, concentrates most of the touristic development, which is the main economic driver of the country, contributing to 18% of the national GDP. Figure 1 – Location of the Cabo Verde archipelago and Sal island as one of its 10 islands. Cabo Verde’s archipelago is an important nesting site for seabirds, with 8 different breeding species. From those, three are endemic species from the archipelago, the Cape Verde Petrel (Pterodroma feae), the Cape Verde Shearwater (Calonectris edwardsii) and the Cape Verde Storm-Petrel (Hydrobates jabejabe). There are also 2 endemic subspecies, the Cape Verde Little Shearwater (Puffinus lherminieri boydi) and the White-faced Storm-Petrel (Pelagodroma marina eadesorum). -

Kapverdische Inseln
KAPVERDISCHE INSELN IT EX M T R A K A R T Mehr entdecken mit MERIAN TopTen 360˚ E FotoTipps für die schönsten Urlaubsmotive Ideen für abwechslungsreiches Reisen mit Kindern live! KAPVERDISCHE INSELN Susanne Lipps ist als Studienreiseleiterin im portugiesischspra- chigen Raum tätig und verfasste Reiseführer über die Kanarischen Inseln, die Azoren und Madeira. Familientipps FotoTipp Barrierefreie Unterkünfte Ziele in der Umgebung Umweltbewusst Reisen Faltkarte Preise für ein Doppelzimmer Preise für ein dreigängiges Menü mit Frühstück: ohne Getränke: €€€€ ab 11 000 ECV €€€€ ab 1400 ECV €€€ ab 8000 ECV €€€ ab 1000 ECV €€ ab 5000 ECV €€ ab 800 ECV € bis 5000 ECV € bis 800 ECV INHALT Willkommen auf den Kapverdischen Inseln 4 MERIAN TopTen 10 Höhepunkte, die Sie sich nicht entgehen lassen sollten 6 MERIAN TopTen 360° Hier finden Sie sich schnell zurecht . 8 MERIAN Tipps 10 Tipps, die Ihnen unbekannte Seiten der Region zeigen 18 Zu Gast auf den Kapverdischen Inseln 22 Übernachten ................................................. 24 Essen und Trinken ........................................... 26 Einkaufen .................................................... 28 Sport und Strände .......................................... 30 Familientipps ................................................ 34 Die Regenzeit sorgt in der Serra da Malagueta ( MERIAN TopTen, S. 64) für üppiges Grün. Unterwegs auf den Kapverdischen Inseln 38 São Vicente, Sal und Boavista . 40 Santo Antão und São Nicolau Santiago und Maio . 54 Fogo und Brava . 70 São Vicente, Santo Antão Sal und Boavista und São Nicolau . 78 Fogo und Brava Santiago und Maio Touren und Ausflüge 100 Kreuz und quer durch Boavista – Entdeckungen in der Einsamkeit . 102 Cidade Velha, Santiago – Romantische Ruinenstadt am Meer . 104 Pistenfahrt durch Sal – Abenteuer abseits der Touristenstrände . 106 Das Tal von Fajã de Água, Brava – Wanderung unter Palmen . -

Cabo Verde Ficha Informativa Ramsar Publicada El 18 Noviembre 2016 Versión Actualizada, Previamente Publicada En 18 Julio 2005
FIR para el Sitio núm. 1576, Lagoa de Rabil, Cabo Verde Ficha Informativa Ramsar Publicada el 18 noviembre 2016 Versión actualizada, previamente publicada en 18 julio 2005 Cabo Verde Lagoa de Rabil Fecha de designación 18 julio 2005 Sitio número 1576 Coordenadas 16°09'N 22°53'57"W Área 113,00 ha https://rsis.ramsar.org/ris/1576 FIR creada por SISR V.1.7 - 1 diciembre 2016 FIR para el Sitio núm. 1576, Lagoa de Rabil, Cabo Verde Código de colores Los campos con fondo sombreado en azul claro se refieren a datos e información necesarios únicamente para las actualizaciones de la FIR. Obsérvese que no está previsto que algunos campos sobre determinados aspectos de la Parte 3, relativos a la Descripción de las Características Ecológicas de la FIR (resaltados en púrpura), se rellenen como parte de una FIR estándar, sino que se incluyen para completar la información con objeto de guardar la coherencia solicitada entre la FIR y el modelo de descripción 'completa' de las características ecológicas aprobado en la Resolución X.15 (2008). En caso de que una Parte Contratante disponga de información relacionada con esos campos (por ejemplo, a partir de un modelo nacional de descripción de las características ecológicas), podrá, si lo desea, aportar información en esos campos adicionales. 1 - Resumen Resumen The Ribeira do Rabil Ramsar Site is made up of the Ribeira do Rabil permanent coastal lagoon, the dune ecosystems at the end of the watercourse and the associated tree and shrub formations. 1. The permanent coastal lagoon contains brackish or salt water, depending on the season (wet or dry).