Oxidation of Hydrogen Sulfide by Quinones
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Orbital-Dependent Redox Potential Regulation of Quinone Derivatives For
RSC Advances View Article Online PAPER View Journal | View Issue Orbital-dependent redox potential regulation of quinone derivatives for electrical energy storage† Cite this: RSC Adv.,2019,9,5164 Zhihui Niu,‡ab Huaxi Wu,‡a Yihua Lu,b Shiyun Xiong,a Xi Zhu,*b Yu Zhao *a and Xiaohong Zhang*a Electrical energy storage in redox flow batteries has received increasing attention. Redox flow batteries using organic compounds, especially quinone-based molecules, as active materials are of particular interest owing to the material sustainability, tailorable redox properties, and environmental friendliness of quinones and their derivatives. In this report, various quinone derivatives were investigated to determine their suitability for applications in organic RFBs. Moreover, the redox potential could be internally Received 14th November 2018 regulated through the tuning of s and p bonding contribution at the redox-active sites. Furthermore, the Accepted 22nd January 2019 binding geometry of some selected quinone derivatives with metal cations was studied. These studies DOI: 10.1039/c8ra09377f provide an alternative strategy to identify and design new quinone molecules with suitable redox rsc.li/rsc-advances potentials for electrical energy storage in organic RFBs. Creative Commons Attribution-NonCommercial 3.0 Unported Licence. Introduction include electrolyte corrosivity, poor redox kinetics, high mate- rial cost, and relatively low efficiency, which prevent their large- The development of renewable energy technologies that are not scale commercialization.9,10 -

Synthesis and Chemistry of Naphthalene Annulated Trienyl Iron Complexes: Potential Anticancer DNA Alkylation Reagents Traci Means University of Arkansas, Fayetteville
Inquiry: The University of Arkansas Undergraduate Research Journal Volume 3 Article 18 Fall 2002 Synthesis and Chemistry of Naphthalene Annulated Trienyl Iron Complexes: Potential Anticancer DNA Alkylation Reagents Traci Means University of Arkansas, Fayetteville Follow this and additional works at: http://scholarworks.uark.edu/inquiry Part of the Medical Biochemistry Commons, and the Organic Chemistry Commons Recommended Citation Means, Traci (2002) "Synthesis and Chemistry of Naphthalene Annulated Trienyl Iron Complexes: Potential Anticancer DNA Alkylation Reagents," Inquiry: The University of Arkansas Undergraduate Research Journal: Vol. 3 , Article 18. Available at: http://scholarworks.uark.edu/inquiry/vol3/iss1/18 This Article is brought to you for free and open access by ScholarWorks@UARK. It has been accepted for inclusion in Inquiry: The nivU ersity of Arkansas Undergraduate Research Journal by an authorized editor of ScholarWorks@UARK. For more information, please contact [email protected], [email protected]. Means: Synthesis and Chemistry of Naphthalene Annulated Trienyl Iron Co 120 INQUIRY Volume 3 2002 SYNTHESIS AND CHEMISTRY OF NAPHTHALENE ANNULATED TRIENYL IRON COMPLEXES: POTENTIAL ANTICANCER DNA ALKYLATION REAGENTS Traci Means Department of Chemistry and Bioc~emistry Fulbright College of Arts and Sctences Mentor: Dr. Neil T. Allison Department of Chemistry and Biochemistry Abstract: is mainly electrophilic in nature, which can also be directly correlated to their toxicological properties. If the alkylation Iron complex chemistry that opens a new door to the process is achieved under mild, ideally biological, conditions, it medicinal and pharmaceutical worlds is the aim ofthis research. could be used in a number ofbiomolecular applications. In fact, Specifically, ortho-quinone methide moieties are intermediates these reactive intermediates have been used as DNA alkylating in several antitumor drugs and have been identified as agents and crosslinkers.1 Compounds that react through quinone bioreductive alkylators ofDNA. -

The Hydrolysis of Carbonyl Sulfide at Low Temperature: a Review
Hindawi Publishing Corporation The Scientific World Journal Volume 2013, Article ID 739501, 8 pages http://dx.doi.org/10.1155/2013/739501 Review Article The Hydrolysis of Carbonyl Sulfide at Low Temperature: A Review Shunzheng Zhao, Honghong Yi, Xiaolong Tang, Shanxue Jiang, Fengyu Gao, Bowen Zhang, Yanran Zuo, and Zhixiang Wang Department of Environmental Engineering, College of Civil and Environmental Engineering, University of Science and Technology Beijing, Beijing 100083, China Correspondence should be addressed to Honghong Yi; [email protected] Received 16 May 2013; Accepted 19 June 2013 Academic Editors: S. Niranjan, L. Wang, and Q. Wang Copyright © 2013 Shunzheng Zhao et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Catalytic hydrolysis technology of carbonyl sulfide (COS) at low temperature was reviewed, including the development of catalysts, reaction kinetics, and reaction mechanism of COS hydrolysis. It was indicated that the catalysts are mainly involved metal oxide and activated carbon. The active ingredients which can load on COS hydrolysis catalyst include alkali metal, alkaline earth metal, transition metal oxides, rare earth metal oxides, mixed metal oxides, and nanometal oxides. The catalytic hydrolysis of COS isa first-order reaction with respect to carbonyl sulfide, while the reaction order of water changes as the reaction conditions change. The controlling steps are also different because the reaction conditions such as concentration of carbonyl sulfide, reaction temperature, water-air ratio, and reaction atmosphere are different. The hydrolysis of carbonyl sulfide is base-catalyzed reaction, and the forceof thebasesitehasanimportanteffectonthehydrolysisofcarbonylsulfide. -

Catechol Ortho-Quinones: the Electrophilic Compounds That Form Depurinating DNA Adducts and Could Initiate Cancer and Other Diseases
Carcinogenesis vol.23 no.6 pp.1071–1077, 2002 Catechol ortho-quinones: the electrophilic compounds that form depurinating DNA adducts and could initiate cancer and other diseases Ercole L.Cavalieri1,3, Kai-Ming Li1, Narayanan Balu1, elevated level of reactive oxygen species, a condition known Muhammad Saeed1, Prabu Devanesan1, as oxidative stress (1,2). As electrophiles, catechol quinones Sheila Higginbotham1, John Zhao2, Michael L.Gross2 and can form covalent adducts with cellular macromolecules, Eleanor G.Rogan1 including DNA (4). These are stable adducts that remain in DNA unless removed by repair and depurinating ones that are 1Eppley Institute for Research in Cancer and Allied Diseases, University of Nebraska Medical Center, 986805 Nebraska Medical Center, Omaha, released from DNA by destabilization of the glycosyl bond. NE 68198-6805 and 2Department of Chemistry, Washington University, Thus, DNA can be damaged by the reactive quinones them- One Brookings Drive, St Louis, MO 63130, USA selves and by reactive oxygen species (hydroxyl radicals) 3To whom correspondence should be addressed (1,4,5). The formation of depurinating adducts by CE quinones Email: [email protected] reacting with DNA may be a major event in the initiation of Catechol estrogens and catecholamines are metabolized to breast and other human cancers (4,5). The depurinating adducts quinones, and the metabolite catechol (1,2-dihydroxyben- are released from DNA, leaving apurinic sites in the DNA zene) of the leukemogenic benzene can also be oxidized to that can generate -
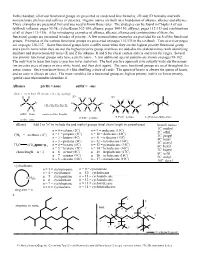
In This Handout, All of Our Functional Groups Are Presented As Condensed Line Formulas, 2D and 3D Formulas and with Nomenclature Prefixes and Suffixes (If Present)
In this handout, all of our functional groups are presented as condensed line formulas, 2D and 3D formulas and with nomenclature prefixes and suffixes (if present). Organic names are built on a foundation of alkanes, alkenes and alkynes. Those examples are presented first and you need to know those rules. The strategies can be found in Chapter 4 of our textbook (alkanes: pages 93-98, cycloalkanes 102-104, alkenes: pages 104-110, alkynes: pages 112-113 and combinations of all of them 113-115). After introducing examples of alkanes, alkenes, alkynes and combinations of them, the functional groups are presented in order of priority. A few nomenclature examples are provided for each of the functional groups. Examples of the various functional groups are presented on pages 115-135 in the textbook. Two overview pages are on pages 136-137. Some functional groups have a suffix name when they are the highest priority functional group and a prefix name when they are not the highest priority group, and these are added to the skeletal names with identifying numbers and stereochemistry terms (E and Z for alkenes, R and S for chiral centers and cis and trans for rings). Several low priority functional groups only have a prefix name. A few additional special patterns are shown on pages 98-102. The only way to learn this topic is practice (over and over). The best practice approach is to actually write out the names (on an extra piece of paper or on a white board, and then do it again). The same functional groups are used throughout the entire course. -

Removal of Hydrogen Sulfide, Benzene and Toluene by a Fluidized Bed Bioreactor
Korean J. Chem. Eng., 23(1), 148-152 (2006) Removal of hydrogen sulfide, benzene and toluene by a fluidized bed bioreactor Kwang Joong Oh†, Ki Chul Cho, Young Hean Choung, Suk Kyong Park*, Sung Ki Cho* and Donguk Kim* Department of Environmental Engineering, Pusan National University, Busan 609-735, Korea *Department of Pharmaceutical Engineering, Inje University, Gimhae, Gyongnam 621-749, Korea (Received 11 May 2005 • accepted 5 September 2005) Abstract−The dominant off gases from publicly owned treatment works include hydrogen sulfide, benzene, and toluene. In this research, hydrogen sulfide oxidized by Bacillus cereus, and benzene with toluene were removed by VOC-degrading microbial consortium. The optimum operating condition of the fluidized bed bioreactor including both microorganisms was 30 oC, pH 6-8, and 150 cm of liquid bed height. The critical loading rate of hydrogen sulfide, benzene and toluene in the bioreactor was about 15 g/m3 h, 10 g/m3 h and 12 g/m3 h, respectively. The fluidized bed bioreactor showed an excellent elimination capacity for 580 hours of continuous operation, and maintained stable removal efficiency at sudden inlet concentration changes. Key words: Hydrogen Sulfide, Benzene, Toluene, Fluidized Bed Bioreactor, Microbial Consortium INTRODUCTION EXPERIMENTAL The major odorous compounds of off-gases from publicly owned 1. Cells 2− treatment works (POTWs) include hydrogen sulfide and VOCs such H2S was oxidized to SO4 by Bacillus cereus which was sepa- as benzene and toluene [Cox and Deshusses, 2001]. Odorous gases rated from closed coal mines in Hwasoon, Korea. Concentration of have been conventionally treated by physical and chemical treat- sulfate in solution was measured by absorbance at 460 nm after reac- ments like absorption, adsorption and catalytic oxidation [Cooper tion with BaCl2·H2O [Cha et al., 1994]. -

Provisional Peer-Reviewed Toxicity Values for Carbonyl Sulfide (Carbon Oxide Sulfide; Casrn 463 58 1)
EPA/690/R-15/002F l Final 9-29-2015 Provisional Peer-Reviewed Toxicity Values for Carbonyl Sulfide (Carbon Oxide Sulfide) (CASRN 463-58-1) Superfund Health Risk Technical Support Center National Center for Environmental Assessment Office of Research and Development U.S. Environmental Protection Agency Cincinnati, OH 45268 AUTHORS, CONTRIBUTORS, AND REVIEWERS CHEMICAL MANAGER John C. Lipscomb, PhD, DABT, Fellow ATS National Center for Environmental Assessment, Cincinnati, OH DRAFT DOCUMENT PREPARED BY SRC, Inc. 7502 Round Pond Road North Syracuse, NY 13212 PRIMARY INTERNAL REVIEWERS Jeff Swartout National Center for Environmental Assessment, Cincinnati, OH This document was externally peer reviewed under contract to Eastern Research Group, Inc. 110 Hartwell Avenue Lexington, MA 02421-3136 Questions regarding the contents of this document may be directed to the U.S. EPA Office of Research and Development’s National Center for Environmental Assessment, Superfund Health Risk Technical Support Center (513-569-7300). ii Carbonyl Sulfide TABLE OF CONTENTS COMMONLY USED ABBREVIATIONS AND ACRONYMS .................................................. iv BACKGROUND .............................................................................................................................1 DISCLAIMERS ...............................................................................................................................1 QUESTIONS REGARDING PPRTVs ............................................................................................1 -

Investigation of Microbial Interactions and Ecosystem
INVESTIGATION OF MICROBIAL INTERACTIONS AND ECOSYSTEM DYNAMICS IN A LOW O2 CYANOBACTERIAL MAT by Alexander A. Voorhies A dissertation submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy (Earth and Environmental Sciences) in The University of Michigan 2014 Doctoral Committee: Assistant Professor Gregory J. Dick, Chair Associate Professor Matthew R. Chapman Assistant Professor Vincent J. Denef Professor Daniel C. Fisher Associate Professor Nathan D. Sheldon © Alexander A. Voorhies 2014 DEDICATION To my wife Hannah ii ACKNOWLEDGEMENTS Funding for the research presented here was provided by the National Science Foundation, the University of Michigan CCMB Pilot Grant, and a Scott Turner research award from the University of Michigan Earth and Environmental Sciences Department. I am grateful for the opportunities to explore my scientific interests this funding has made possible. I would like to acknowledge my co-authors and collaborators, who offered advice, guidance and immeasurable assistance throughout this process. Gregory J. Dick, Bopi Biddanda, Scott T. Kendall, Sunit Jain, Daniel N. Marcus, Stephen C. Nold and Nathan D. Sheldon are co- authors on CHAPTER II, which was published in Geobiology in 2012; Gregory J. Dick was a co-author on CHAPTER III, which is in preparation for publication; and Gregory J. Dick, Sarah D. Eisenlord, Daniel N. Marcus, Melissa B. Duhaime, Bopaiah A. Biddanda and James D Cavalcoli are co-authors on Chapter IV, which is in preparation for publication. I thank my committee members: Matt Chapman, Nathan Sheldon, Vincent Denef and Dan Fisher. Their input and guidance throughout my graduate studies has kept me on track and made significant enhancements to this dissertation. -

The Respiratory Chain & Oxidative Phosphorylation
ch12.qxd 2/13/2003 2:46 PM Page 92 The Respiratory Chain & Oxidative Phosphorylation 12 Peter A. Mayes, PhD, DSc, & Kathleen M. Botham, PhD, DSc BIOMEDICAL IMPORTANCE trapping the liberated free energy as high-energy phos- phate, and the enzymes of β-oxidation and of the citric Aerobic organisms are able to capture a far greater pro- acid cycle (Chapters 22 and 16) that produce most of portion of the available free energy of respiratory sub- the reducing equivalents. strates than anaerobic organisms. Most of this takes place inside mitochondria, which have been termed the “powerhouses” of the cell. Respiration is coupled to the Components of the Respiratory Chain generation of the high-energy intermediate, ATP, by Are Arranged in Order of Increasing oxidative phosphorylation, and the chemiosmotic Redox Potential theory offers insight into how this is accomplished. A number of drugs (eg, amobarbital) and poisons (eg, Hydrogen and electrons flow through the respiratory chain (Figure 12–3) through a redox span of 1.1 V cyanide, carbon monoxide) inhibit oxidative phos- + phorylation, usually with fatal consequences. Several in- from NAD /NADH to O2/2H2O (Table 11–1). The herited defects of mitochondria involving components respiratory chain consists of a number of redox carriers of the respiratory chain and oxidative phosphorylation that proceed from the NAD-linked dehydrogenase sys- have been reported. Patients present with myopathy tems, through flavoproteins and cytochromes, to mole- and encephalopathy and often have lactic acidosis. cular oxygen. Not all substrates are linked to the respi- ratory chain through NAD-specific dehydrogenases; some, because their redox potentials are more positive SPECIFIC ENZYMES ACT AS MARKERS (eg, fumarate/succinate; Table 11–1), are linked di- rectly to flavoprotein dehydrogenases, which in turn are OF COMPARTMENTS SEPARATED BY linked to the cytochromes of the respiratory chain (Fig- THE MITOCHONDRIAL MEMBRANES ure 12–4). -

Investigating PAH Relative Reactivity Using Congener Profiles, Quinone Measurements and Back Trajectories
Open Access Discussion Paper | Discussion Paper | Discussion Paper | Discussion Paper | Atmos. Chem. Phys. Discuss., 13, 25741–25768, 2013 Atmospheric www.atmos-chem-phys-discuss.net/13/25741/2013/ Chemistry doi:10.5194/acpd-13-25741-2013 © Author(s) 2013. CC Attribution 3.0 License. and Physics Discussions This discussion paper is/has been under review for the journal Atmospheric Chemistry and Physics (ACP). Please refer to the corresponding final paper in ACP if available. Investigating PAH relative reactivity using congener profiles, quinone measurements and back trajectories M. S. Alam1, J. M. Delgado-Saborit1, C. Stark1, and R. M. Harrison1,2 1Division of Environmental Health & Risk Management School of Geography, Earth & Environmental Sciences University of Birmingham Edgbaston, Birmingham, B15 2TT, UK 2Department of Environmental Sciences/Center of Excellence in Environmental Studies, King Abdulaziz University, Jeddah, 21589, Saudi Arabia Received: 22 July 2013 – Accepted: 12 September 2013 – Published: 8 October 2013 Correspondence to: R. M. Harrison ([email protected]) Published by Copernicus Publications on behalf of the European Geosciences Union. 25741 Discussion Paper | Discussion Paper | Discussion Paper | Discussion Paper | Abstract Vapour and particle-associated concentrations of 15 polycyclic aromatic hydrocarbons (PAH) and 11 PAH quinones have been measured in winter and summer campaigns at the rural site, Weybourne in eastern England. Concentrations of individual PAH are 20– 5 140 times smaller than average concentrations at an English urban site. The concen- trations of PAH are greatest in air masses originating from southern England relative to those from Scandinavia and the North Atlantic, while quinone to parent PAH ratios show an inverse behaviour, being highest in the more aged North Atlantic polar air masses. -

Converting Copper Sulfide to Copper with Surface Sulfur For
ARTICLE https://doi.org/10.1038/s41467-021-24059-y OPEN Converting copper sulfide to copper with surface sulfur for electrocatalytic alkyne semi- hydrogenation with water ✉ Yongmeng Wu 1,3, Cuibo Liu 1,3, Changhong Wang 1,3, Yifu Yu 1, Yanmei Shi 1 & Bin Zhang 1,2 Electrocatalytic alkyne semi-hydrogenation to alkenes with water as the hydrogen source using a low-cost noble-metal-free catalyst is highly desirable but challenging because of their 1234567890():,; over-hydrogenation to undesired alkanes. Here, we propose that an ideal catalyst should have the appropriate binding energy with active atomic hydrogen (H*) from water electrolysis and a weaker adsorption with an alkene, thus promoting alkyne semi-hydrogenation and avoiding over-hydrogenation. So, surface sulfur-doped and -adsorbed low-coordinated copper nano- wire sponges are designedly synthesized via in situ electroreduction of copper sulfide and enable electrocatalytic alkyne semi-hydrogenation with over 99% selectivity using water as the hydrogen source, outperforming a copper counterpart without surface sulfur. Sulfur 2− + 2− + anion-hydrated cation (S -K (H2O)n) networks between the surface adsorbed S and K in the KOH electrolyte boost the production of active H* from water electrolysis. And the trace doping of sulfur weakens the alkene adsorption, avoiding over-hydrogenation. Our catalyst also shows wide substrate scopes, up to 99% alkenes selectivity, good reducible groups compatibility, and easily synthesized deuterated alkenes, highlighting the promising potential of this method. 1 Department of Chemistry, Institute of Molecular Plus, School of Science, Tianjin University, Tianjin, China. 2 Tianjin Key Laboratory of Molecular Optoelectronic Science, Collaborative Innovation Center of Chemical Science and Engineering, Tianjin, China. -

Boom Clay Natural Organic Matter
EXTERNAL REPORT SCK•CEN-ER-206 12/CBr/P-37 Boom Clay natural organic matter Status Report 2011 Christophe Bruggeman and Mieke De Craen SCK•CEN Contract: CO-90-08-2214-00 NIRAS/ONDRAF contract: CCHO 2009- 0940000 Research Plan Geosynthesis June, 2012 SCK•CEN RDD Boeretang 200 BE-2400 Mol Belgium EXTERNAL REPORT OF THE BELGIAN NUCLEAR RESEARCH CENTRE SCK•CEN-ER-206 12/CBr/P-37 Boom Clay natural organic matter Status Report 2011 Christophe Bruggeman and Mieke De Craen SCK•CEN Contract: CO-90-08-2214-00 NIRAS/ONDRAF contract: CCHO 2009- 0940000 Research Plan Geosynthesis June, 2012 Status: Unclassified ISSN 1782-2335 SCK•CEN Boeretang 200 BE-2400 Mol Belgium © SCK•CEN Studiecentrum voor Kernenergie Centre d’étude de l’énergie Nucléaire Boeretang 200 BE-2400 Mol Belgium Phone +32 14 33 21 11 Fax +32 14 31 50 21 http://www.sckcen.be Contact: Knowledge Centre [email protected] COPYRIGHT RULES All property rights and copyright are reserved to SCK•CEN. In case of a contractual arrangement with SCK•CEN, the use of this information by a Third Party, or for any purpose other than for which it is intended on the basis of the contract, is not authorized. With respect to any unauthorized use, SCK•CEN makes no representation or warranty, expressed or implied, and assumes no liability as to the completeness, accuracy or usefulness of the information contained in this document, or that its use may not infringe privately owned rights. SCK•CEN, Studiecentrum voor Kernenergie/Centre d'Etude de l'Energie Nucléaire Stichting van Openbaar Nut – Fondation d'Utilité Publique ‐ Foundation of Public Utility Registered Office: Avenue Herrmann Debroux 40 – BE‐1160 BRUSSEL Operational Office: Boeretang 200 – BE‐2400 MOL Table of Contents Abstract .....................................................................................................................................