Screening Pyridine Derivatives Against Human Hydrogen Sulfide-Synthesizing Enzymes by Orthogonal Methods
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Hydrolysis of Carbonyl Sulfide at Low Temperature: a Review
Hindawi Publishing Corporation The Scientific World Journal Volume 2013, Article ID 739501, 8 pages http://dx.doi.org/10.1155/2013/739501 Review Article The Hydrolysis of Carbonyl Sulfide at Low Temperature: A Review Shunzheng Zhao, Honghong Yi, Xiaolong Tang, Shanxue Jiang, Fengyu Gao, Bowen Zhang, Yanran Zuo, and Zhixiang Wang Department of Environmental Engineering, College of Civil and Environmental Engineering, University of Science and Technology Beijing, Beijing 100083, China Correspondence should be addressed to Honghong Yi; [email protected] Received 16 May 2013; Accepted 19 June 2013 Academic Editors: S. Niranjan, L. Wang, and Q. Wang Copyright © 2013 Shunzheng Zhao et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Catalytic hydrolysis technology of carbonyl sulfide (COS) at low temperature was reviewed, including the development of catalysts, reaction kinetics, and reaction mechanism of COS hydrolysis. It was indicated that the catalysts are mainly involved metal oxide and activated carbon. The active ingredients which can load on COS hydrolysis catalyst include alkali metal, alkaline earth metal, transition metal oxides, rare earth metal oxides, mixed metal oxides, and nanometal oxides. The catalytic hydrolysis of COS isa first-order reaction with respect to carbonyl sulfide, while the reaction order of water changes as the reaction conditions change. The controlling steps are also different because the reaction conditions such as concentration of carbonyl sulfide, reaction temperature, water-air ratio, and reaction atmosphere are different. The hydrolysis of carbonyl sulfide is base-catalyzed reaction, and the forceof thebasesitehasanimportanteffectonthehydrolysisofcarbonylsulfide. -
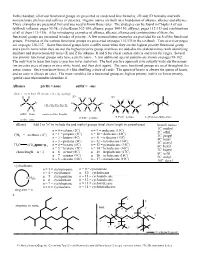
In This Handout, All of Our Functional Groups Are Presented As Condensed Line Formulas, 2D and 3D Formulas and with Nomenclature Prefixes and Suffixes (If Present)
In this handout, all of our functional groups are presented as condensed line formulas, 2D and 3D formulas and with nomenclature prefixes and suffixes (if present). Organic names are built on a foundation of alkanes, alkenes and alkynes. Those examples are presented first and you need to know those rules. The strategies can be found in Chapter 4 of our textbook (alkanes: pages 93-98, cycloalkanes 102-104, alkenes: pages 104-110, alkynes: pages 112-113 and combinations of all of them 113-115). After introducing examples of alkanes, alkenes, alkynes and combinations of them, the functional groups are presented in order of priority. A few nomenclature examples are provided for each of the functional groups. Examples of the various functional groups are presented on pages 115-135 in the textbook. Two overview pages are on pages 136-137. Some functional groups have a suffix name when they are the highest priority functional group and a prefix name when they are not the highest priority group, and these are added to the skeletal names with identifying numbers and stereochemistry terms (E and Z for alkenes, R and S for chiral centers and cis and trans for rings). Several low priority functional groups only have a prefix name. A few additional special patterns are shown on pages 98-102. The only way to learn this topic is practice (over and over). The best practice approach is to actually write out the names (on an extra piece of paper or on a white board, and then do it again). The same functional groups are used throughout the entire course. -

Removal of Hydrogen Sulfide, Benzene and Toluene by a Fluidized Bed Bioreactor
Korean J. Chem. Eng., 23(1), 148-152 (2006) Removal of hydrogen sulfide, benzene and toluene by a fluidized bed bioreactor Kwang Joong Oh†, Ki Chul Cho, Young Hean Choung, Suk Kyong Park*, Sung Ki Cho* and Donguk Kim* Department of Environmental Engineering, Pusan National University, Busan 609-735, Korea *Department of Pharmaceutical Engineering, Inje University, Gimhae, Gyongnam 621-749, Korea (Received 11 May 2005 • accepted 5 September 2005) Abstract−The dominant off gases from publicly owned treatment works include hydrogen sulfide, benzene, and toluene. In this research, hydrogen sulfide oxidized by Bacillus cereus, and benzene with toluene were removed by VOC-degrading microbial consortium. The optimum operating condition of the fluidized bed bioreactor including both microorganisms was 30 oC, pH 6-8, and 150 cm of liquid bed height. The critical loading rate of hydrogen sulfide, benzene and toluene in the bioreactor was about 15 g/m3 h, 10 g/m3 h and 12 g/m3 h, respectively. The fluidized bed bioreactor showed an excellent elimination capacity for 580 hours of continuous operation, and maintained stable removal efficiency at sudden inlet concentration changes. Key words: Hydrogen Sulfide, Benzene, Toluene, Fluidized Bed Bioreactor, Microbial Consortium INTRODUCTION EXPERIMENTAL The major odorous compounds of off-gases from publicly owned 1. Cells 2− treatment works (POTWs) include hydrogen sulfide and VOCs such H2S was oxidized to SO4 by Bacillus cereus which was sepa- as benzene and toluene [Cox and Deshusses, 2001]. Odorous gases rated from closed coal mines in Hwasoon, Korea. Concentration of have been conventionally treated by physical and chemical treat- sulfate in solution was measured by absorbance at 460 nm after reac- ments like absorption, adsorption and catalytic oxidation [Cooper tion with BaCl2·H2O [Cha et al., 1994]. -

Provisional Peer-Reviewed Toxicity Values for Carbonyl Sulfide (Carbon Oxide Sulfide; Casrn 463 58 1)
EPA/690/R-15/002F l Final 9-29-2015 Provisional Peer-Reviewed Toxicity Values for Carbonyl Sulfide (Carbon Oxide Sulfide) (CASRN 463-58-1) Superfund Health Risk Technical Support Center National Center for Environmental Assessment Office of Research and Development U.S. Environmental Protection Agency Cincinnati, OH 45268 AUTHORS, CONTRIBUTORS, AND REVIEWERS CHEMICAL MANAGER John C. Lipscomb, PhD, DABT, Fellow ATS National Center for Environmental Assessment, Cincinnati, OH DRAFT DOCUMENT PREPARED BY SRC, Inc. 7502 Round Pond Road North Syracuse, NY 13212 PRIMARY INTERNAL REVIEWERS Jeff Swartout National Center for Environmental Assessment, Cincinnati, OH This document was externally peer reviewed under contract to Eastern Research Group, Inc. 110 Hartwell Avenue Lexington, MA 02421-3136 Questions regarding the contents of this document may be directed to the U.S. EPA Office of Research and Development’s National Center for Environmental Assessment, Superfund Health Risk Technical Support Center (513-569-7300). ii Carbonyl Sulfide TABLE OF CONTENTS COMMONLY USED ABBREVIATIONS AND ACRONYMS .................................................. iv BACKGROUND .............................................................................................................................1 DISCLAIMERS ...............................................................................................................................1 QUESTIONS REGARDING PPRTVs ............................................................................................1 -

Converting Copper Sulfide to Copper with Surface Sulfur For
ARTICLE https://doi.org/10.1038/s41467-021-24059-y OPEN Converting copper sulfide to copper with surface sulfur for electrocatalytic alkyne semi- hydrogenation with water ✉ Yongmeng Wu 1,3, Cuibo Liu 1,3, Changhong Wang 1,3, Yifu Yu 1, Yanmei Shi 1 & Bin Zhang 1,2 Electrocatalytic alkyne semi-hydrogenation to alkenes with water as the hydrogen source using a low-cost noble-metal-free catalyst is highly desirable but challenging because of their 1234567890():,; over-hydrogenation to undesired alkanes. Here, we propose that an ideal catalyst should have the appropriate binding energy with active atomic hydrogen (H*) from water electrolysis and a weaker adsorption with an alkene, thus promoting alkyne semi-hydrogenation and avoiding over-hydrogenation. So, surface sulfur-doped and -adsorbed low-coordinated copper nano- wire sponges are designedly synthesized via in situ electroreduction of copper sulfide and enable electrocatalytic alkyne semi-hydrogenation with over 99% selectivity using water as the hydrogen source, outperforming a copper counterpart without surface sulfur. Sulfur 2− + 2− + anion-hydrated cation (S -K (H2O)n) networks between the surface adsorbed S and K in the KOH electrolyte boost the production of active H* from water electrolysis. And the trace doping of sulfur weakens the alkene adsorption, avoiding over-hydrogenation. Our catalyst also shows wide substrate scopes, up to 99% alkenes selectivity, good reducible groups compatibility, and easily synthesized deuterated alkenes, highlighting the promising potential of this method. 1 Department of Chemistry, Institute of Molecular Plus, School of Science, Tianjin University, Tianjin, China. 2 Tianjin Key Laboratory of Molecular Optoelectronic Science, Collaborative Innovation Center of Chemical Science and Engineering, Tianjin, China. -

Vinyl Alkyl Sulfides Via Hydrozirconation of Terminal Alkynes
J. Serb. Chem. Soc. 69(3)175–178(2004) UDC 678.745+542.913 JSCS – 3142 Original scientific paper Stereoselective synthesis of (E)-vinyl alkyl sulfides via hydrozirconation of terminal alkynes PING ZHONG*1,2 and XIAN HUANG2 1Department of Chemistry, Wenzhou Normal College, Wenzhou 325027, P. R. China and 2Department of Chemistry, Zhejiang University, Hangzhou 310028, P. R. China (e-mail: [email protected]) (Received 15 July, revised 3 November 2003) h5 Abstract: Terminal alkynes react with Cp2Zr(H)Cl (Cp = -C5H5)togiveorgano- zirconium complexes, which are trapped with alkylsulfenyl chlorides to afford (E)-vinyl alkyl sulfides in good yield. Keywords: alkyne, sulfide, alkylsulfenyl chloride, hydrozirconation, synthesis. INTRODUCTION Vinyl sulfides have attracted special atention as important intermediates in vari- ous synthetic transformations,1,2 and their stereospecific syntheses has been attemp- ted.3,4 To our knowledge, there are only a few stereoselective synthesis of 1-alkenyl sulfides reported. These methods include the reduction of vinyl sulfoxides by EtMgBr/CI,5 the cross-coupling reaction of 1-alkenyl halides with metal thioalkoxides in the presence of a transition-metal catalyst,6 as well as the reaction of b-sulfinium vi- nyl boron compounds with catecholborane cross-coupling on organic halides7 in the presence of the catalyst PdCl2(dppf). The two first methods require stereodefined vinylsulfoxides and vinyl halides, which cannot always be obtained with high ste- reo-selectivity. The corresponding vinyl halides are also needed in the third one and a phase-transfer catalyst with a longer reaction time. A convenient stereoselecitve method for the synthesis of (E)-vinyl sulfides will be described in this paper. -

(12) United States Patent (10) Patent No.: US 9,079,857 B2 Hani Et Al
US0090798.57B2 (12) United States Patent (10) Patent No.: US 9,079,857 B2 Hani et al. (45) Date of Patent: Jul. 14, 2015 (54) PROCESS FOR PREPARING ALKALI METAL 3,297,556 A 1/1967 Boudakian PYRTHONE AND ITS POLYVALENT 3,590,035 A 6, 1971 Damico METAL COMPLEXES FROM PYRONE 3,700,676 A 10/1972 Damico OXDE 3,773,770 A 1 1/1973 Damico 3,899,495 A 8, 1975 Beschke et al. (71) Applicant: Arch Chemicals, Inc., Atlanta, GA (US) 4,080,329 A 3/1978 Muntwyler 4,396,766 A 8/1983 Farmer, Jr. et al. (72) Inventors: Rahim Hani, Alpharetta, GA (US); is: 38. R John J. Jardas, Rochester, NY (US); 5.536.376 A T/1996 N al Richard Dumas, East Haven, CT (US); - - w amaguchi et al. David Lei, Alpharetta,s GA (US)s s 5,869,678 A 2f1999 Schiesslet al. FOREIGN PATENT DOCUMENTS (73) Assignee: Arch Chemicals, Inc., Atlanta, GA (US) DE 2208007 9, 1973 (*) Notice: Subject to any disclaimer, the term of this DE 2.717325 11, 1977 patent is extended or adjusted under 35 JP S 58O88362 5, 1983 U.S.C. 154(b) by 5 days. JP S 58152867 9, 1983 JP S 59112968 6, 1984 JP HO13O8255 12/1989 (21) Appl. No.: 13/838,470 KR 2002003 1981 5, 2002 (22) Filed: Mar 15, 2013 OTHER PUBLICATIONS (65) Prior Publication Data International Search Report and Written Opinion, PCT/US2013/ US 2013/0261309 A1 Oct. 3, 2013 034882, mailed May 15, 2013. O O J. Chinese Medicine Industry, 21(7), 317, 1990. -

Survey of Key Functional Groups Overview Overview Overview
11/2/2009 Overview Survey of Key Functional Groups Ch 8.1 Chem 201 – Functional Group Survey 11/2/2009 Overview Overview Chem 201 – Functional Group Survey 11/2/2009 Chem 201 – Functional Group Survey 11/2/2009 1 11/2/2009 Concerns Functional Group Names Getting overwhelmed Alcohol Strange new formulas Strange new names Strange new reagents Alkyl halide, Haloalkane Many new relationships leads to brain meltdown !!! Info builds ONE step at a Thiol time Ch 8 surveys FG Names, drawings, properties No new reactions (almost) Ether Sulfide Chem 201 – Functional Group Survey 11/2/2009 Chem 201 – Functional Group Survey 11/2/2009 RX Names Common or Substitutive (IUPAC)? Special Compounds Use Common Names Alkyl halide Haloalkane Haloforms Alkyl group attached Halogen substitutent methylene chloride to Halogen attached to Alkane chloroform ()(dichloromethane) bromoform Methyl chloride Chloromethane iodoform Ethyl bromide Bromoethane Useful solvents Special compounds carbon tetrachloride - hydrophobic Special groups - moderately polar carbon tetrabromide - volatile methylene, allyl, vinyl, benzyl but all linked to health problems Special history allyl bromide these slides will be posted Chem 201 – Functional Group Survey 11/2/2009 Chem 201 – Functional Group Survey 11/2/2009 2 11/2/2009 Give “haloalkane” (IUPAC) names Give “haloalkane” (IUPAC) names 2- fluoropropane cis-121,2-dibromocyc lo hexane Chem 201 – Functional Group Survey 11/2/2009 Chem 201 – Functional Group Survey 11/2/2009 ROR’ Names ROR’ Names Common & Substitutive -

Oxidation of Hydrogen Sulfide by Quinones
International Journal of Molecular Sciences Article Oxidation of Hydrogen Sulfide by Quinones: How Polyphenols Initiate Their Cytoprotective Effects Kenneth R. Olson 1,2,* , Yan Gao 1 and Karl D. Straub 3,4 1 Indiana University School of Medicine—South Bend Center, South Bend, IN 46617, USA; [email protected] 2 Department of Biological Sciences, University of Notre Dame, Notre Dame, IN 46556, USA 3 Central Arkansas Veteran’s Healthcare System, Little Rock, AR 72205, USA; [email protected] 4 Departments of Medicine and Biochemistry, University of Arkansas for Medical Sciences, Little Rock, AR 72202, USA * Correspondence: [email protected]; Tel.: +1-(574)-631-7560; Fax: +1-(574)-631-7821 Abstract: We have shown that autoxidized polyphenolic nutraceuticals oxidize H2S to polysulfides and thiosulfate and this may convey their cytoprotective effects. Polyphenol reactivity is largely at- tributed to the B ring, which is usually a form of hydroxyquinone (HQ). Here, we examine the effects of HQs on sulfur metabolism using H2S- and polysulfide-specific fluorophores (AzMC and SSP4, respectively) and thiosulfate sensitive silver nanoparticles (AgNP). In buffer, 1,4-dihydroxybenzene (1,4-DB), 1,4-benzoquinone (1,4-BQ), pyrogallol (PG) and gallic acid (GA) oxidized H2S to polysul- fides and thiosulfate, whereas 1,2-DB, 1,3-DB, 1,2-dihydroxy,3,4-benzoquinone and shikimic acid did not. In addition, 1,4-DB, 1,4-BQ, PG and GA also increased polysulfide production in HEK293 cells. In buffer, H2S oxidation by 1,4-DB was oxygen-dependent, partially inhibited by tempol and trolox, and absorbance spectra were consistent with redox cycling between HQ autoxidation and H2S-mediated reduction. -

The Selective Oxidation of Sulfides to Sulfoxides Or Sulfones with Hydrogen Peroxide Catalyzed by a Dendritic Phosphomolybdate H
catalysts Article The Selective Oxidation of Sulfides to Sulfoxides or Sulfones with Hydrogen Peroxide Catalyzed by a Dendritic Phosphomolybdate Hybrid Qiao-Lin Tong y, Zhan-Fang Fan y, Jian-Wen Yang, Qi Li, Yi-Xuan Chen, Mao-Sheng Cheng and Yang Liu * Key Laboratory of Structure-Based Drug Design & Discovery of Ministry of Education, School of Pharmaceutical Engineering, Shenyang Pharmaceutical University, Shenyang 110016, China; [email protected] (Q.-L.T.); [email protected] (Z.-F.F.); [email protected] (J.-W.Y.); [email protected] (Q.L.); [email protected] (Y.-X.C.); [email protected] (M.-S.C.) * Correspondence: [email protected] Qiao-Lin Tong and Zhan-Fang Fan contributed equally to this work. y Received: 6 September 2019; Accepted: 20 September 2019; Published: 22 September 2019 Abstract: The oxidation of sulfides to their corresponding sulfoxides or sulfones has been achieved using a low-cost poly(amidoamine) with a first-generation coupled phosphomolybdate hybrid as the catalyst and aqueous hydrogen peroxide as the oxidant. The reusability of the catalyst was revealed in extensive experiments. The practice of this method in the preparation of a smart drug Modafinil has proved its good applicability. Keywords: dendritic phosphomolybdate hybrid; sulfides; selective oxidation; hydrogen peroxide; reusability 1. Introduction Many biologically and chemically active molecules are constructed from sulfoxides and sulfones [1–5]. The oxidation of sulfides is a fundamental reaction as one of the most straightforward methods to afford sulfoxides and sulfones [6]. Many reagents, including peracids and halogen derivatives, are used in the common approaches of sulfoxidation reactions [7,8]. -

Carbon Disulfide
Chapter 5.4 Carbon disulfide General Description Carbon disulfide (CS2) in its pure form is a colourless, volatile and in-flammable liquid with a sweet aromatic odour. The technical product is a yellowish liquid with a disagreeable odour. Sources Carbon disulfide is used in large quantities as an industrial chemical for the production of viscose rayon fibres. In this technological process, for every kilogram of viscose produced, about 20-30 g of carbon disulfide and 4-6 g of hydrogen sulfide are emitted (1). Additional release of carbon disulfide, carbonyl sulfide and hydrogen sulfide takes place from coal gasification plants; data on the total emission from these plants are not available. The ventilation discharge from viscose plants can reach several millions of m3 per hour, with a carbon disulfide content varying from 20 to 240 mg/m3, which represents a total emission of 15-40 tonnes of carbon disulfide daily (2). Exposure to carbon disulfide is mostly confined to those engaged in technological processes in the viscose industry. However, the general population living near viscose plants may also be exposed to carbon disulfide emissions. Occurrence in air The primary source of carbon disulfide in the environment is emission from viscose plants, around which environmental pollution is especially great. A scientific review of Soviet literature indicates values ranging from 0.01 to 0.21 mg/m3 around viscose plants (2). A recent Austrian study reports that concentrations of 0.05 ppm (157 μg/m3) were often exceeded in the vicinity of viscose plants, even at a distance of several kilometres, and concentrations close to the plants could be 5-10 times higher. -

Thiol Reactive Probes and Chemosensors
Sensors 2012, 12, 15907-15946; doi:10.3390/s121115907 OPEN ACCESS sensors ISSN 1424-8220 www.mdpi.com/journal/sensors Review Thiol Reactive Probes and Chemosensors Hanjing Peng 1, Weixuan Chen 1, Yunfeng Cheng 1, Lovemore Hakuna 2, Robert Strongin 2 and Binghe Wang 1,* 1 Department of Chemistry and the Center for Diagnostics and Therapeutics, Center for Biotechnology and Drug Design, Georgia State University, Atlanta, GA 30302, USA; E-Mails: [email protected] (H.P.); [email protected] (W.C.); [email protected] (Y.C.) 2 Department of Chemistry, Portland State University, Portland, OR 97207, USA; E-Mails: [email protected] (L.H.); [email protected] (R.S.) * Author to whom correspondence should be addressed; E-Mail: [email protected]; Tel.: +1-404-413-5545; Fax: +1-404-413-5543. Received: 24 September 2012; in revised form: 12 November 2012 / Accepted: 13 November 2012 / Published: 19 November 2012 Abstract: Thiols are important molecules in the environment and in biological processes. Cysteine (Cys), homocysteine (Hcy), glutathione (GSH) and hydrogen sulfide (H2S) play critical roles in a variety of physiological and pathological processes. The selective detection of thiols using reaction-based probes and sensors is very important in basic research and in disease diagnosis. This review focuses on the design of fluorescent and colorimetric probes and sensors for thiol detection. Thiol detection methods include probes and labeling agents based on nucleophilic addition and substitution, Michael addition, disulfide bond or Se-N bond cleavage, metal-sulfur interactions and more. Probes for H2S are based on nucleophilic cyclization, reduction and metal sulfide formation.