Influence of Acetylcholine Esterase Inhibitors and Memantine, Clinically
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Relief of Hypersensitivity After Nerve Injury from Systemic Donepezil
Saranya devi Relief of Hypersensitivity after Nerve Injury from ALN Systemic Donepezil Involves Spinal Cholinergic and γ-Aminobutyric Acid Mechanisms Spinal ACh and GABA Mechanisms for Donepezil Analgesia Masafumi Kimura, M.D.,* Ken-ichiro Hayashida, D.V.M., Ph.D.,† James C. Eisenach, M.D.,‡ KIMURA ET AL. Shigeru Saito M.D.,§ Hideaki Obata, M.D.‖ XX ABSTRACT What We Already Know about This Topic • Donepezil increases central nervous system acetylcholine lev- Downloaded from http://pubs.asahq.org/anesthesiology/article-pdf/118/1/173/259568/0000542-201301000-00030.pdf by guest on 28 September 2021 10.1097/ALN.0b013e318277a81c Background: Evoking spinal release of acetylcholine (ACh) els and is used for treatment of dementia produces antinociception in normal animals and reduces • Increases in spinal acetylcholine levels have been implicated in the relief of neuropathic pain hypersensitivity after nerve injury, and some studies suggest January that ACh-mediated analgesia relies on γ-aminobutyric acid (GABA)-ergic signaling in the spinal cord. In this study, the authors tested the spinal mechanisms underlying the anti- What This Article Tells Us That Is New 118 hypersensitivity effects of donepezil, a central nervous sys- • Donepezil increased spinal acetylcholine levels and γ- tem–penetrating cholinesterase inhibitor, in a rat model of aminobutyric acid levels to reduce nociceptive responses after nerve injury and represents a potential therapeutic pathway to neuropathic pain. reduce pain after nerve injury Methods: Male Sprague-Dawley rats were anesthetized, and L5 spinal nerve ligation was performed unilaterally. With- A antagonist; and CGP 35348 (30 μg), a γ-aminobutyric drawal threshold to a paw pressure test was measured before acid receptor type B antagonist. -

Tor Amendment Tagoor Laboratories Private Limited
ToR Amendment Tagoor Laboratories Private Limited ANNEXURE – I Details of Products as per Granted ToR and Amendment Required Production Capacity S. Ton /Month Therapeutic Product Name No As per Amendment category granted ToR Required 1 Abacavir sulfate 2.0 2.0 Anti HIV 2 Amitriptyline 5.0 10.0 Antidepressant 3 Atrovastatin Calcium 5.0 5.0 Hypercholesterolemia 4 Bupropion 5.0 5.0 Anti depressant 5 carisoprodol 2.0 2.0 Muscle Relaxant 6 Clopidogrelbisulfate 5.0 5.0 Antithrombotic 7 Cyclobenzaprine HCl 5.0 5.0 Muscle relaxant 8 Cyproheptadine HCl 5.0 10.0 Anti allergic 9 Desloratadine 5.0 5.0 Antihistamine 10 Domperidone 20.0 30.0 Anti emetic 11 Domperidone maleate 2.0 2.0 Anti emetic 12 Donepezil HCl 1.0 1.0 Alzheimer’s disease 13 Ebastine 5.0 5.0 Anti allergic 14 Esomeprazole Sodium 3.0 3.0 Anti ulcerative Esomeprazole Magnesium 3.0 3.0 Anti ulcerative 15 trihydrate 16 Fexofenadine Hydrochloride 6.0 15.0 Anti histamine 17 Haloperidol 2.0 2.0 Antipsychotic 18 Itopride Hydrochloride 2.0 2.0 Antispasmodics 19 Itraconazole 10.0 10.0 Antifungal 20 Ketrolac Tromethane 2.0 2.0 Anti Inflammatory 21 Lansoprazole 10.0 10.0 Ant ulcerative 22 Loperamide Hydrochloride 10.0 10.0 Anti diarrhea agent 23 Losartan Potassium 6.0 15.0 Anti Hypertensive 24 Nebivolol HCl 2.0 2.0 Anti Hypertensive 25 Nortriptyline HCl 2.0 2.0 Anti depressant 26 Omeprazole 40.0 40.0 Anti ulcerative 27 Omeprazole Sodium 2.0 2.0 Ant ulcerative Omeprazole Magnesium 2.0 2.0 Ant ulcerative 28 Dihydrate 29 Oxatomide 1.0 1.0 Antihistamine Pantoprazole Sodium Sesqui 10.0 20.0 Ant ulcerative 30 Hydrate 31 Pimozide 2.0 2.0 Antipsychotic 32 Pregabalin 2.0 2.0 Epileptic 33 Quetiapine Hemifumarate 2.0 2.0 Antipsychotic Prepared By Rightsource Industrial Solutions Pvt. -
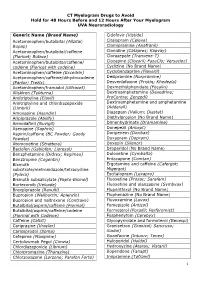
CT Myelogram Drugs to Avoid Hold for 48 Hours Before and 12 Hours After Your Myelogram UVA Neuroradiology
CT Myelogram Drugs to Avoid Hold for 48 Hours Before and 12 Hours After Your Myelogram UVA Neuroradiology Generic Name (Brand Name) Cidofovir (Vistide) Acetaminophen/butalbital (Allzital; Citalopram (Celexa) Bupap) Clomipramine (Anafranil) Acetaminophen/butalbital/caffeine Clonidine (Catapres; Kapvay) (Fioricet; Butace) Clorazepate (Tranxene-T) Acetaminophen/butalbital/caffeine/ Clozapine (Clozaril; FazaClo; Versacloz) codeine (Fioricet with codeine) Cyclizine (No Brand Name) Acetaminophen/caffeine (Excedrin) Cyclobenzaprine (Flexeril) Acetaminophen/caffeine/dihydrocodeine Desipramine (Norpramine) (Panlor; Trezix) Desvenlafaxine (Pristiq; Khedezla) Acetaminophen/tramadol (Ultracet) Dexmethylphenidate (Focalin) Aliskiren (Tekturna) Dextroamphetamine (Dexedrine; Amitriptyline (Elavil) ProCentra; Zenzedi) Amitriptyline and chlordiazepoxide Dextroamphetamine and amphetamine (Limbril) (Adderall) Amoxapine (Asendin) Diazepam (Valium; Diastat) Aripiprazole (Abilify) Diethylpropion (No Brand Name) Armodafinil (Nuvigil) Dimenhydrinate (Dramamine) Asenapine (Saphris) Donepezil (Aricept) Aspirin/caffeine (BC Powder; Goody Doripenem (Doribax) Powder) Doxapram (Dopram) Atomoxetine (Strattera) Doxepin (Silenor) Baclofen (Gablofen; Lioresal) Droperidol (No Brand Name) Benzphetamine (Didrex; Regimex) Duloxetine (Cymbalta) Benztropine (Cogentin) Entacapone (Comtan) Bismuth Ergotamine and caffeine (Cafergot; subcitrate/metronidazole/tetracycline Migergot) (Pylera) Escitalopram (Lexapro) Bismuth subsalicylate (Pepto-Bismol) Fluoxetine (Prozac; Sarafem) -

Retrospective Observation of Liver Function Parameters for 101 Patients Using Herbal Drugs for One Month
31 2 (2010 3 ) J Korean Oriental Med 2010;31(2):149-157 Original article 김동민1, 김회권2, 조성연3, 김용석1, 남상수1 1경희대학교 한의과대학 침구학교실, 2가평군 보건소, 3경희대학교 강남경희한방병원 침구과 Retrospective Observation of Liver Function Parameters for 101 Patients Using Herbal Drugs for One Month Dong-Min Kim 1, Hyee-Kwon Kim 2, Seong-Yeun Cho 3, Yong-Suk Kim 1, Sang-Soo Nam 1 1Dept. of Acupuncture & Moxibustion, College of Oriental Medicine, Kyung Hee University 2Gapyeong Public Health Center 3Dept. of Acupuncture & Moxibustion, Kangnam KyungHee Korean Hospital, Kyung Hee University Objective: The purpose of this study was to investigate the level of safety on liver functions when Korean herbal medicine was taken internally. Method: 101 inpatients who took Korean herbal medicine were enrolled and liver function test (aspartic aminotransferase, alanine aminotransferase, alkaline phosphatase, total bilirubin, gamma glutamyltranspeptidase, albumin, lactate dehydrogenase) was performed on admission and 1 month later. Results: In 101 patients, alkaline phosphatase and gamma glutamyltranspeptidase decreased significantly compared with the value taken on admission (p<0.05) but aspartic aminotransferase, alanine aminotransferase, total bilirubin, albumin, and lactate dehydrogenase were not significantly changed (p>0.05). In the patients who took Scutellaria baicalensis (n=34), alkaline phosphatase decreased and albumin increased significantly (p<0.05). Among the patients who took Atractylodes macrocephala (n=29), alkaline phosphatase decreased significantly (p<0.05). In the patients who took Glycyrrhiza uralensis or Paeonia lactiflora , liver function parameters were not significantly changed (p>0.05). On admission 11 patients had abnormal liver function and 2 patients had liver injury while 7 patients had abnormal liver function and 2 patients showed liver injury 1 month later. -

Overselling Donepezil (ARICEPD and Exploiting Patients V1ith I\Lzheimers Disease: Why Isn't the FDA Stopping These Ads?
PUBLIC CITIZEN HEALTH RESEARCH GROUP SIDNEY M. WOLFE, M.D., EDITOR March 2004 + VOL. 20, N0 . 3 Overselling Donepezil (ARICEPD and Exploiting Patients v1ith i\lzheimers Disease: Why Isn't the FDA Stopping These Ads? n the 1999 Edition of Worst Pills, patients. The ads proclaim that If this conclusion were based on a Best Pills, we urged patients not patients with Alzheimer's disease randomized, placebo-controlled trial I to use donepezil (ARICEPT), who "persistently" use donepezil are (RCT) (the scientific gold standard for made by Pfizer/ Esai, because of a able to stay in the community and medical research) wherein 50 percent combination of adverse effects such avoid going into a nursing home for of the patients got donepezil and the as nausea, diarrhea and vomiting almost two years more than patients other 50 percent of exactly the same along with serious questions about who have only limited treatment or kinds of patients were randomly clinically significant effectiveness. no treatment with the dmg (see assigned to be given a placebo, it We quoted the prestigious Medical advertisement on page 2 for an would be a major public health Letter, an independent evaluation for example). The implication is star breakthrough and a cause for cele physicians of newly approved dmgs, tling. The families and other friends bration. Unfortunately, it is not based which stated "There is no evidence who are caring for patients with on such a study and is not any cause that use of... donepezil... leads to Alzheimer's disease will have a for celebration. substantial functional improvement substantial amount of extra time to The study cited as the basis for this or prevents the progression of the have their loved ones in the commu seemingly remarkable improvement disease." Since then, Pfizer/ Esai has nity before they have to go into a is flawed. -

Drugs Affectin the Autonomic Nervous System
Fundamentals of Medical Pharmacology Paterson Public Schools Written by Néstor Collazo, Ph.D. Jonathan Hodges, M.D. Tatiana Mikhaelovsky, M.D. for Health and Related Professions (H.A.R.P.) Academy March 2007 Course Description This fourth year course is designed to give students in the Health and Related Professions (H.A.R.P.) Academy a general and coherent explanation of the science of pharmacology in terms of its basic concepts and principles. Students will learn the properties and interactions between chemical agents (drugs) and living organisms for the rational and safe use of drugs in the control, prevention, and therapy of human disease. The emphasis will be on the fundamental concepts as they apply to the actions of most prototype drugs. In order to exemplify important underlying principles, many of the agents in current use will be singled out for fuller discussion. The course will include the following topics: ¾ The History of Pharmacology ¾ Terminology Used in Pharmacology ¾ Drug Action on Living Organisms ¾ Principles of Pharmacokinetics ¾ Dose-Response Relationships ¾ Time-Response Relationships ¾ Human Variability: Factors that will modify effects of drugs on individuals ¾ Effects of Drugs Attributable to Varying Modes of Administration ¾ Drug Toxicity ¾ Pharmacologic Aspects of Drug Abuse and Drug Dependence Pre-requisites Students must have completed successfully the following courses: Biology, Chemistry, Anatomy and Physiology, Algebra I and II Credits: 5 credits Basic Principles of Drug Action Introduction to Pharmacology a. Basic Mechanisms of Drug Actions b. Dose-response relationships c. Drug absorption d. Biotransformation of Drugs e. Pharmacokinetics f. Factors Affecting Drug Distribution g. Drug Allergy and Pharmacogenetics h. -

Potential Herb–Drug Interactions in the Management of Age-Related Cognitive Dysfunction
pharmaceutics Review Potential Herb–Drug Interactions in the Management of Age-Related Cognitive Dysfunction Maria D. Auxtero 1, Susana Chalante 1,Mário R. Abade 1 , Rui Jorge 1,2,3 and Ana I. Fernandes 1,* 1 CiiEM, Interdisciplinary Research Centre Egas Moniz, Instituto Universitário Egas Moniz, Quinta da Granja, Monte de Caparica, 2829-511 Caparica, Portugal; [email protected] (M.D.A.); [email protected] (S.C.); [email protected] (M.R.A.); [email protected] (R.J.) 2 Polytechnic Institute of Santarém, School of Agriculture, Quinta do Galinheiro, 2001-904 Santarém, Portugal 3 CIEQV, Life Quality Research Centre, IPSantarém/IPLeiria, Avenida Dr. Mário Soares, 110, 2040-413 Rio Maior, Portugal * Correspondence: [email protected]; Tel.: +35-12-1294-6823 Abstract: Late-life mild cognitive impairment and dementia represent a significant burden on health- care systems and a unique challenge to medicine due to the currently limited treatment options. Plant phytochemicals have been considered in alternative, or complementary, prevention and treat- ment strategies. Herbals are consumed as such, or as food supplements, whose consumption has recently increased. However, these products are not exempt from adverse effects and pharmaco- logical interactions, presenting a special risk in aged, polymedicated individuals. Understanding pharmacokinetic and pharmacodynamic interactions is warranted to avoid undesirable adverse drug reactions, which may result in unwanted side-effects or therapeutic failure. The present study reviews the potential interactions between selected bioactive compounds (170) used by seniors for cognitive enhancement and representative drugs of 10 pharmacotherapeutic classes commonly prescribed to the middle-aged adults, often multimorbid and polymedicated, to anticipate and prevent risks arising from their co-administration. -

Donepezil, a Drug for Alzheimer’S Disease, Promotes Oligodendrocyte
www.nature.com/aps ARTICLE Donepezil, a drug for Alzheimer’s disease, promotes oligodendrocyte generation and remyelination Xue Cui1, Yu-e Guo2,3, Jia-hui Fang1, Chang-jie Shi1, Na Suo2,3, Ru Zhang1 and Xin Xie1,2,4 Myelin sheaths play important roles in neuronal functions. In the central nervous system (CNS), the myelin is formed by oligodendrocytes (OLs), which are differentiated from oligodendrocyte precursor cells (OPCs). In CNS demyelinating disorders such as multiple sclerosis (MS), the myelin sheaths are damaged and the remyelination process is hindered. Small molecule drugs that promote OPC to OL differentiation and remyelination may provide a new way to treat these demyelinating diseases. Here we report that donepezil, an acetylcholinesterase inhibitor (AChEI) developed for the treatment of Alzheimer’s disease (AD), significantly promotes OPC to OL differentiation. Interestingly, other AChEIs, including huperzine A, rivastigmine, and tacrine, have no such effect, indicating that donepezil’s effect in promoting OPC differentiation is not dependent on the inhibition of AChE. Donepezil also facilitates the formation of myelin sheaths in OPC–DRG neuron co-culture. More interestingly, donepezil also promotes the repair of the myelin sheaths in vivo and provides significant therapeutic effect in a cuprizone-mediated demyelination animal model. Donepezil is a drug that has been used to treat AD safely for many years; our findings suggest that it might be repurposed to treat CNS demyelinating diseases such as MS by promoting OPC to OL differentiation and remyelination. Keywords: donepezil; aricept; oligodendrocyte; oligodendrocyte progenitor cell; differentiation; myelin sheaths; remyelination; CNS demyelinating disorders; multiple sclerosis Acta Pharmacologica Sinica (2019) 40:1386–1393; https://doi.org/10.1038/s41401-018-0206-4 INTRODUCTION mitochondria function, has been found to accelerate OL matura- Oligodendrocytes (OLs), an important component of neuroglia, tion in culture and increase remyelination in rodents [11, 12]. -

In-Vitro Evaluation of Selected Egyptian Traditional Herbal Medicines for Treatment of Alzheimer Disease
Ali et al. BMC Complementary and Alternative Medicine 2013, 13:121 http://www.biomedcentral.com/1472-6882/13/121 RESEARCH ARTICLE Open Access In-vitro evaluation of selected Egyptian traditional herbal medicines for treatment of alzheimer disease Shereen K Ali1, Ahmed R Hamed1,2, Maha M Soltan1,2,UsamaMHegazy3, Esameldin E Elgorashi4, Ibrahim A El-Garf5 and Ahmed A Hussein6* Abstract Background: Egyptians recognized the healing power of herbs and used them in their medicinal formulations. Nowadays, “Attarin” drug shops and the public use mainly the Unani medicinal system for treatment of their health problems including improvement of memory and old age related diseases. Numerous medicinal plants have been described in old literature of Arabic traditional medicine for treatment of Alzheimer’s disease (AD) (or to strengthen memory). Methods: In this study, some of these plants were evaluated against three different preliminary bioassays related to AD to explore the possible way of their bio-interaction. Twenty three selected plants were extracted with methanol and screened in vitro against acetylcholinesterase (AChE) and cycloxygenase-1 (COX-1) enzymes. In addition, anti-oxidant activity using DPPH was determined. Results: Of the tested plant extracts; Adhatoda vasica and Peganum harmala showed inhibitory effect on AChE at IC50 294 μg/ml and 68 μg/ml respectively. Moreover, A. vasica interacted reversibly with the enzyme while P. harmala showed irreversible inhibition. Ferula assafoetida (IC50 3.2 μg/ml), Syzygium aromaticum (34.9 μg/ml) and Zingiber officinalis (33.6 μg/ml) showed activity against COX-1 enzyme. Potent radical scavenging activity was demonstrated by three plant extracts Terminalia chebula (EC50 2.2 μg/ml), T. -

Acetylcholinesterase: the “Hub” for Neurodegenerative Diseases And
Review biomolecules Acetylcholinesterase: The “Hub” for NeurodegenerativeReview Diseases and Chemical Weapons Acetylcholinesterase: The “Hub” for Convention Neurodegenerative Diseases and Chemical WeaponsSamir F. de A. Cavalcante Convention 1,2,3,*, Alessandro B. C. Simas 2,*, Marcos C. Barcellos 1, Victor G. M. de Oliveira 1, Roberto B. Sousa 1, Paulo A. de M. Cabral 1 and Kamil Kuča 3,*and Tanos C. C. França 3,4,* Samir F. de A. Cavalcante 1,2,3,* , Alessandro B. C. Simas 2,*, Marcos C. Barcellos 1, Victor1 Institute G. M. ofde Chemical, Oliveira Biological,1, Roberto Radiological B. Sousa and1, Paulo Nuclear A. Defense de M. Cabral (IDQBRN),1, Kamil Brazilian Kuˇca Army3,* and TanosTechnological C. C. França Center3,4,* (CTEx), Avenida das Américas 28705, Rio de Janeiro 23020-470, Brazil; [email protected] (M.C.B.); [email protected] (V.G.M.d.O.); [email protected] 1 Institute of Chemical, Biological, Radiological and Nuclear Defense (IDQBRN), Brazilian Army (R.B.S.); [email protected] (P.A.d.M.C.) Technological Center (CTEx), Avenida das Américas 28705, Rio de Janeiro 23020-470, Brazil; 2 [email protected] Mors Institute of Research (M.C.B.); on Natural [email protected] Products (IPPN), Federal (V.G.M.d.O.); University of Rio de Janeiro (UFRJ), CCS,[email protected] Bloco H, Rio de Janeiro (R.B.S.); 21941-902, [email protected] Brazil (P.A.d.M.C.) 32 DepartmentWalter Mors of Institute Chemistry, of Research Faculty of on Science, Natural Un Productsiversity (IPPN), -

Cholinesterase Inhibitors of Cholinergic Activity in Rat Isolated Stomach
Proceedings of the British Pharmacological Society at http://www.pA2online.org/abstracts/Vol4Issue2abst121P.pdf POTENTIATION BY CHOLINESTERASE INHIBITORS OF CHOLINERGIC ACTIVITY IN RAT ISOLATED STOMACH Emma M. Jarvie, Sandrine Ociepka, Selim Cellek, Jason D. Brown & Gareth J. Sanger Neurology and Gastrointestinal CEDD, GlaxoSmithKline, Harlow, Essex, CM19 5AW, UK. Cholinesterase (ChE) inhibitors increase gastrointestinal motility, but related side effects are common. These may reach a 53 % (nausea) or 12 % (diarrhoea) incidence with the acetyl-ChE (AChE) and butyryl-ChE (BuChE) inhibitor neostigmine and possibly lower with the selective AChE inhibitor donepezil (Imbimbo, 2001). However, the AChE inhibitor and dopamine D2 receptor antagonist itopride seems well tolerated (e.g. Sawant et al., 2004). We compared their AChE inhibitory activity and their gastric prokinetic-like activity in vitro, by measuring changes in cholinergically-mediated contraction, evoked by electrical field stimulation (EFS; Bassil et al., 2006). Longitudinal forestomach preparations from adult, fed male CD rats (200 - 250 g) were suspended between two ring electrodes in Krebs’ buffer (5% CO2/ 95% O2; pH 7.4; 37°C) under tension for isometric recording. TTX (1 µM)- and scopolamine (10 µM)- sensitive contractions were evoked by EFS (25 V, 2.5 Hz, 0.5 ms pulse duration, for 10 s every 1 min). Compounds were added cumulatively (at least 10 min contact) to the bathing solution. AChE activity was measured over 5 min, using AChE from human erythrocytes incubated with 5,5'-dithiobis-(2-nitro-benzoic acid) before addition of acetylthiocholine (Ellman et al., 1961). Data are means ± standard error of the mean; n = animals used (stomach) or number of experiments (erythrocytes) per compound. -

Evaluation of the Effects of Donepezil in Psychotic Disorders Using Swiss Albino Mice
International Journal of Basic & Clinical Pharmacology Tanwani H et al. Int J Basic Clin Pharmacol. 2020 Apr;9(4):533-537 http:// www.ijbcp.com pISSN 2319-2003 | eISSN 2279-0780 DOI: http://dx.doi.org/10.18203/2319-2003.ijbcp20201051 Original Research Article Evaluation of the effects of donepezil in psychotic disorders using Swiss albino mice Hemant Tanwani1, Ritesh Churihar2*, Sameer Pandit1 1Department of Pharmacology, MGM Medical College, Indore, Madhya Pradesh, India 2Department of Pharmacology, Govt. Medical College, Ratlam , Madhya Pradesh, India Received: 27 January 2020 Accepted: 10 February 2020 *Correspondence: Dr. Ritesh Churihar, Email: [email protected] Copyright: © the author(s), publisher and licensee Medip Academy. This is an open-access article distributed under the terms of the Creative Commons Attribution Non-Commercial License, which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited. ABSTRACT Background: Schizophrenia as a psychotic disorder is currently treated by various antipsychotic drugs. A large group of patients still remain resistant to the treatment and present in the form of residual cognitive deficits. Donepezil has been advocated at various conferences and seminars for using it in schizophrenia patients. Donepezil is currently approved drug for Alzheimer's disease to improve cognition. Hence, we have tried to assess its role for psychotic models induced by methylphenidate in mice. Methods: Methylphenidate 5 mg/kg was given by intraperitoneal (i.p) route to induce psychosis in Swiss albino mice (n=6). Donepezil was given alone in a dose of 1 mg/kg and in combination with low dose haloperidol 0.1 mg/kg and groups were compared with haloperidol 0.2 mg/kg.