Pretreatment and Prophylaxis Against Nerve Agent Poisoning: Are Undesirable Behavioral Side Effects Unavoidable?
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Rheumatoid Arthritis Induced by Botulinum Toxin Type A: a Case Report and Review of the Literature
Open Access Austin Journal of Orthopedics & Rheumatology Case Report Rheumatoid Arthritis Induced by Botulinum Toxin Type A: A Case Report and Review of the Literature Yanyan G1,2#, Yupeng L1#, Yuanyuan L1,3, Fangfang Z1,3 and Meiying W1* Abstract 1 Department of Rheumatology and Immunology, Introduction: Botulinum Toxin Type A (BoNT/A) is a bacterial toxin Shenzhen Second People’s Hospital, The First Affiliated commonly used in cosmetic therapy. Although there has been a great deal of Hospital of Shenzhen University, Shenzhen, China clinical and basic research on the potential therapeutic applications of botulinum 2Department of Nephrology, Peking University Shenzhen toxin there are few reports on its clinical toxicity and side effects. Hospital, Shenzhen, China 3Department of Rheumatology and Immunology, Peking Patient Concerns: A previously healthy 26-year-old woman developed University Shenzhen Hospital, Shenzhen, China joint pain and redness in her right toe, swelling in the posterior left foot and #Contributed equally to this paper the interphalangeal joint of the right index finger, occasional shoulder pain, and morning stiffness for 30 minutes daily, 6 months after BoNT/A injection. *Corresponding author: Meiying Wang, Department of Rheumatology and Immunology, Shenzhen Second Diagnosis: Laboratory testing showed elevated Rheumatoid Factor (RF), People’s Hospital, The First Affiliated Hospital of anti-cyclic citrullinated peptide (anti-CCP), C-Reactive Protein (CRP), Erythrocyte Shenzhen University, Shenzhen 518035, China Sedimentation Rate (ESR). Doppler ultrasound examination of the right hand and both feet showed synovial hyperplasia of the right wrist, right second Received: April 21, 2021; Accepted: May 15, 2021; proximal interphalangeal joint, both ankles, and right first metatarsophalangeal Published: May 22, 2021 joint, as well as bony erosion in a left intertarsal joint. -

Galantamine Potentiates the Neuroprotective Effect of Memantine Against NMDA-Induced Excitotoxicity Joao~ P
Galantamine potentiates the neuroprotective effect of memantine against NMDA-induced excitotoxicity Joao~ P. Lopes1, Glauco Tarozzo1, Angelo Reggiani1, Daniele Piomelli1,2 & Andrea Cavalli1,3 1D3 – Drug Discovery and Development Department, Istituto Italiano di Tecnologia, Via Morego, 16163, Genova, Italy 2Departments of Anatomy and Neurobiology and Biological Chemistry, University of California, Irvine, CA, 92697-4621 3Department of Pharmacy and Biotechnologies, Alma Mater Studiorum, Bologna University, Via Belmeloro, 40126, Bologna, Italy Keywords Abstract Alzheimer’s disease, drug combination, N NMDA neurotoxicity, NR2B, The combination of memantine, an -methyl-D-aspartate (NMDA) receptor polypharmacology, primary cortical neurons antagonist, with an acetylcholinesterase inhibitor (AChEI) is the current stan- dard of care in Alzheimer’s disease (AD). Galantamine, an AChEI currently Correspondence marketed for the treatment of AD, exerts memory-enhancing and neuroprotec- Andrea Cavalli, D3 – Drug Discovery and tive effects via activation of nicotinic acetylcholine receptors (nAChRs). Here, Development Department, Istituto Italiano we investigated the neuroprotective properties of galantamine in primary cul- di Tecnologia – Via Morego, 30, 16163 tures of rat cortical neurons when given alone or in combination with meman- Genova, Italy. Tel: +39 010 71781530; Fax: +39 010 tine. In agreement with previous findings, we found that memantine was fully 71781228; E-mail: [email protected] effective in reversing NMDA toxicity at concentrations of 2.5 and 5 lmol/L. Galantamine also completely reversed NMDA toxicity at a concentration of Funding Information 5 lmol/L. The a7 and a4b2 nAChR antagonists, methyllycaconitine, and dihy- No funding information provided. dro-b-erythroidine blocked the neuroprotective effect of galantamine, demon- strating the involvement of nAChRs. -

Reference List of Drugs with Potential Anticholinergic Effects 1, 2, 3, 4, 5
ANTICHOLINERGICS: Reference List of Drugs with Potential Anticholinergic Effects 1, 2, 3, 4, 5 J Bareham BSP © www.RxFiles.ca Aug 2021 WHENEVER POSSIBLE, AVOID DRUGS WITH MODERATE TO HIGH ANTICHOLINERGIC ACTIVITY IN OLDER ADULTS (>65 YEARS OF AGE) Low Anticholinergic Activity; Moderate/High Anticholinergic Activity -B in combo Beers Antibiotics Antiparkinsonian Cardiovascular Agents Immunosuppressants ampicillin *ALL AVAILABLE AS amantadine SYMMETREL atenolol TENORMIN azaTHIOprine IMURAN cefOXitin GENERIC benztropine mesylate COGENTIN captopril CAPOTEN cyclosporine NEORAL clindamycin bromocriptine PARLODEL chlorthalidone GENERIC ONLY hydrocortisone CORTEF gentamicin (Oint & Sol’n NIHB covered) carbidopa/levodopa SINEMET digoxin LANOXIN, TOLOXIN methylprednisolone MEDROL piperacillin entacapone COMTAN dilTIAZem CARDIZEM, TIAZAC prednisone WINPRED dipyridamole PERSANTINE, ethopropazine PARSITAN vancomycin phenelzine NARDIL AGGRENOX disopyramide RYTHMODAN Muscle Relaxants pramipexole MIRAPEX Antidepressants baclofen LIORESAL ( on intrathecal only) procyclidine KEMADRIN furosemide LASIX amitriptyline ELAVIL cyclobenzaprine FLEXERIL selegiline ELDEPRYL hydrALAZINE APRESOLINE clomiPRAMINE ANAFRANIL isosorbide ISORDIL methocarbamol ROBAXIN OTC trihexyphenidyl ARTANE desipramine NORPRAMIN metoprolol LOPRESOR orphenadrine NORFLEX OTC doxepin >6mg SINEQUAN Antipsychotics NIFEdipine ADALAT tiZANidine ZANAFLEX A imipramine TOFRANIL quiNIDine GENERIC ONLY C ARIPiprazole ABILIFY & MAINTENA -

Pyridostigmine in the Treatment of Postural
Pyridostigmine in the Treatment of Postural Orthostatic Tachycardia: A Single-Center Experience KHALIL KANJWAL, M.D.,* BEVERLY KARABIN, PH.D.,* MUJEEB SHEIKH, M.D.,* LAWRENCE ELMER, M.D., PH.D.,† YOUSUF KANJWAL, M.D.,* BILAL SAEED, M.D.,* and BLAIR P. GRUBB, M.D.* From the *Electrophysiology Section, Division of Cardiology, Department of Medicine, The University of Toledo, Toledo, Ohio; and †Department of Neurology, The University of Toledo College of Medicine, Health Science Campus, Toledo, Ohio Background: The long-term efficacy of pyridostigmine, a reversible acetyl cholinesterase inhibitor, in the treatment of postural orthostatic tachycardia syndrome (POTS) patients remains unclear. We report our retrospective, single-center, long-term experience regarding the efficacy and adverse effect profile of pyridostigmine in the treatment of POTS patients. Methods: This retrospective study included an extensive review of electronic charts and data collection in regards to patient demographics, orthostatic parameters, side-effect profile, subjective response to therapy, as well as laboratory studies recorded at each follow-up visit to our institution’s Syncope and Autonomic Disorders Center. The response to pyridostigmine therapy was considered successful if patient had both symptom relief in addition to an objective response in orthostatic hemodynamic parameters (heart rate [HR] and blood pressure). Three hundred patients with POTS were screened for evaluation in this study. Of these 300, 203 patients with POTS who received pyridostigmine therapy were reviewed. Of these 203 patients, 168 were able to tolerate the medication after careful dose titration. The mean follow- up duration in this group of patients was 12 ± 3 (9–15) months. Pyridostigmine improved symptoms of orthostatic intolerance in 88 of 203 (43%) of total patients or 88 of 172 (51%) who were able to tolerate the drug. -

Relief of Hypersensitivity After Nerve Injury from Systemic Donepezil
Saranya devi Relief of Hypersensitivity after Nerve Injury from ALN Systemic Donepezil Involves Spinal Cholinergic and γ-Aminobutyric Acid Mechanisms Spinal ACh and GABA Mechanisms for Donepezil Analgesia Masafumi Kimura, M.D.,* Ken-ichiro Hayashida, D.V.M., Ph.D.,† James C. Eisenach, M.D.,‡ KIMURA ET AL. Shigeru Saito M.D.,§ Hideaki Obata, M.D.‖ XX ABSTRACT What We Already Know about This Topic • Donepezil increases central nervous system acetylcholine lev- Downloaded from http://pubs.asahq.org/anesthesiology/article-pdf/118/1/173/259568/0000542-201301000-00030.pdf by guest on 28 September 2021 10.1097/ALN.0b013e318277a81c Background: Evoking spinal release of acetylcholine (ACh) els and is used for treatment of dementia produces antinociception in normal animals and reduces • Increases in spinal acetylcholine levels have been implicated in the relief of neuropathic pain hypersensitivity after nerve injury, and some studies suggest January that ACh-mediated analgesia relies on γ-aminobutyric acid (GABA)-ergic signaling in the spinal cord. In this study, the authors tested the spinal mechanisms underlying the anti- What This Article Tells Us That Is New 118 hypersensitivity effects of donepezil, a central nervous sys- • Donepezil increased spinal acetylcholine levels and γ- tem–penetrating cholinesterase inhibitor, in a rat model of aminobutyric acid levels to reduce nociceptive responses after nerve injury and represents a potential therapeutic pathway to neuropathic pain. reduce pain after nerve injury Methods: Male Sprague-Dawley rats were anesthetized, and L5 spinal nerve ligation was performed unilaterally. With- A antagonist; and CGP 35348 (30 μg), a γ-aminobutyric drawal threshold to a paw pressure test was measured before acid receptor type B antagonist. -

Corticosterone and Pyridostigmine/DEET Exposure
HHS Public Access Author manuscript Author ManuscriptAuthor Manuscript Author Neurotoxicology Manuscript Author . Author Manuscript Author manuscript; available in PMC 2019 May 24. Published in final edited form as: Neurotoxicology. 2019 January ; 70: 26–32. doi:10.1016/j.neuro.2018.10.006. Corticosterone and pyridostigmine/DEET exposure attenuate peripheral cytokine expression: Supporting a dominant role for neuroinflammation in a mouse model of Gulf War Illness Lindsay T. Michalovicza, Alicia R. Lockera, Kimberly A. Kellya, Julie V. Millera, Zachary Barnesb,c, Mary Ann Fletcherb,c, Diane B. Millera, Nancy G. Klimasb,c, Mariana Morrisb, Stephen M. Lasleyd, and James P. O’Callaghana,* aHealth Effects Laboratory Division, Centers for Disease Control and Prevention, National Institute for Occupational Safety and Health, Morgantown, WV, USA bInstitute for Neuro-Immune Medicine, Nova Southeastern University, Ft. Lauderdale, FL, USA cMiami Veterans Affairs Medical Center, Miami, FL, USA dDepartment of Cancer Biology & Pharmacology, University of Illinois College of Medicine at Peoria, Peoria, IL, USA Abstract Gulf War Illness (GWI) is a chronic multi-symptom disorder experienced by as many as a third of the veterans of the 1991 Gulf War; the constellation of “sickness behavior” symptoms observed in ill veterans is suggestive of a neuroimmune involvement. Various chemical exposures and conditions in theater have been implicated in the etiology of the illness. Previously, we found that GW-related organophosphates (OPs), such as the sarin surrogate, DFP, and chlorpyrifos, cause neuroinflammation. The combination of these exposures with exogenous corticosterone (CORT), mimicking high physiological stress, exacerbates the observed neuroinflammation. The potential relationship between the effects of OPs and CORT on the brain versus inflammation in the periphery has not been explored. -

Pharmacy and Poisons (Third and Fourth Schedule Amendment) Order 2017
Q UO N T FA R U T A F E BERMUDA PHARMACY AND POISONS (THIRD AND FOURTH SCHEDULE AMENDMENT) ORDER 2017 BR 111 / 2017 The Minister responsible for health, in exercise of the power conferred by section 48A(1) of the Pharmacy and Poisons Act 1979, makes the following Order: Citation 1 This Order may be cited as the Pharmacy and Poisons (Third and Fourth Schedule Amendment) Order 2017. Repeals and replaces the Third and Fourth Schedule of the Pharmacy and Poisons Act 1979 2 The Third and Fourth Schedules to the Pharmacy and Poisons Act 1979 are repealed and replaced with— “THIRD SCHEDULE (Sections 25(6); 27(1))) DRUGS OBTAINABLE ONLY ON PRESCRIPTION EXCEPT WHERE SPECIFIED IN THE FOURTH SCHEDULE (PART I AND PART II) Note: The following annotations used in this Schedule have the following meanings: md (maximum dose) i.e. the maximum quantity of the substance contained in the amount of a medicinal product which is recommended to be taken or administered at any one time. 1 PHARMACY AND POISONS (THIRD AND FOURTH SCHEDULE AMENDMENT) ORDER 2017 mdd (maximum daily dose) i.e. the maximum quantity of the substance that is contained in the amount of a medicinal product which is recommended to be taken or administered in any period of 24 hours. mg milligram ms (maximum strength) i.e. either or, if so specified, both of the following: (a) the maximum quantity of the substance by weight or volume that is contained in the dosage unit of a medicinal product; or (b) the maximum percentage of the substance contained in a medicinal product calculated in terms of w/w, w/v, v/w, or v/v, as appropriate. -

Tor Amendment Tagoor Laboratories Private Limited
ToR Amendment Tagoor Laboratories Private Limited ANNEXURE – I Details of Products as per Granted ToR and Amendment Required Production Capacity S. Ton /Month Therapeutic Product Name No As per Amendment category granted ToR Required 1 Abacavir sulfate 2.0 2.0 Anti HIV 2 Amitriptyline 5.0 10.0 Antidepressant 3 Atrovastatin Calcium 5.0 5.0 Hypercholesterolemia 4 Bupropion 5.0 5.0 Anti depressant 5 carisoprodol 2.0 2.0 Muscle Relaxant 6 Clopidogrelbisulfate 5.0 5.0 Antithrombotic 7 Cyclobenzaprine HCl 5.0 5.0 Muscle relaxant 8 Cyproheptadine HCl 5.0 10.0 Anti allergic 9 Desloratadine 5.0 5.0 Antihistamine 10 Domperidone 20.0 30.0 Anti emetic 11 Domperidone maleate 2.0 2.0 Anti emetic 12 Donepezil HCl 1.0 1.0 Alzheimer’s disease 13 Ebastine 5.0 5.0 Anti allergic 14 Esomeprazole Sodium 3.0 3.0 Anti ulcerative Esomeprazole Magnesium 3.0 3.0 Anti ulcerative 15 trihydrate 16 Fexofenadine Hydrochloride 6.0 15.0 Anti histamine 17 Haloperidol 2.0 2.0 Antipsychotic 18 Itopride Hydrochloride 2.0 2.0 Antispasmodics 19 Itraconazole 10.0 10.0 Antifungal 20 Ketrolac Tromethane 2.0 2.0 Anti Inflammatory 21 Lansoprazole 10.0 10.0 Ant ulcerative 22 Loperamide Hydrochloride 10.0 10.0 Anti diarrhea agent 23 Losartan Potassium 6.0 15.0 Anti Hypertensive 24 Nebivolol HCl 2.0 2.0 Anti Hypertensive 25 Nortriptyline HCl 2.0 2.0 Anti depressant 26 Omeprazole 40.0 40.0 Anti ulcerative 27 Omeprazole Sodium 2.0 2.0 Ant ulcerative Omeprazole Magnesium 2.0 2.0 Ant ulcerative 28 Dihydrate 29 Oxatomide 1.0 1.0 Antihistamine Pantoprazole Sodium Sesqui 10.0 20.0 Ant ulcerative 30 Hydrate 31 Pimozide 2.0 2.0 Antipsychotic 32 Pregabalin 2.0 2.0 Epileptic 33 Quetiapine Hemifumarate 2.0 2.0 Antipsychotic Prepared By Rightsource Industrial Solutions Pvt. -

Title 16. Crimes and Offenses Chapter 13. Controlled Substances Article 1
TITLE 16. CRIMES AND OFFENSES CHAPTER 13. CONTROLLED SUBSTANCES ARTICLE 1. GENERAL PROVISIONS § 16-13-1. Drug related objects (a) As used in this Code section, the term: (1) "Controlled substance" shall have the same meaning as defined in Article 2 of this chapter, relating to controlled substances. For the purposes of this Code section, the term "controlled substance" shall include marijuana as defined by paragraph (16) of Code Section 16-13-21. (2) "Dangerous drug" shall have the same meaning as defined in Article 3 of this chapter, relating to dangerous drugs. (3) "Drug related object" means any machine, instrument, tool, equipment, contrivance, or device which an average person would reasonably conclude is intended to be used for one or more of the following purposes: (A) To introduce into the human body any dangerous drug or controlled substance under circumstances in violation of the laws of this state; (B) To enhance the effect on the human body of any dangerous drug or controlled substance under circumstances in violation of the laws of this state; (C) To conceal any quantity of any dangerous drug or controlled substance under circumstances in violation of the laws of this state; or (D) To test the strength, effectiveness, or purity of any dangerous drug or controlled substance under circumstances in violation of the laws of this state. (4) "Knowingly" means having general knowledge that a machine, instrument, tool, item of equipment, contrivance, or device is a drug related object or having reasonable grounds to believe that any such object is or may, to an average person, appear to be a drug related object. -
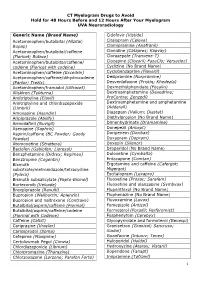
CT Myelogram Drugs to Avoid Hold for 48 Hours Before and 12 Hours After Your Myelogram UVA Neuroradiology
CT Myelogram Drugs to Avoid Hold for 48 Hours Before and 12 Hours After Your Myelogram UVA Neuroradiology Generic Name (Brand Name) Cidofovir (Vistide) Acetaminophen/butalbital (Allzital; Citalopram (Celexa) Bupap) Clomipramine (Anafranil) Acetaminophen/butalbital/caffeine Clonidine (Catapres; Kapvay) (Fioricet; Butace) Clorazepate (Tranxene-T) Acetaminophen/butalbital/caffeine/ Clozapine (Clozaril; FazaClo; Versacloz) codeine (Fioricet with codeine) Cyclizine (No Brand Name) Acetaminophen/caffeine (Excedrin) Cyclobenzaprine (Flexeril) Acetaminophen/caffeine/dihydrocodeine Desipramine (Norpramine) (Panlor; Trezix) Desvenlafaxine (Pristiq; Khedezla) Acetaminophen/tramadol (Ultracet) Dexmethylphenidate (Focalin) Aliskiren (Tekturna) Dextroamphetamine (Dexedrine; Amitriptyline (Elavil) ProCentra; Zenzedi) Amitriptyline and chlordiazepoxide Dextroamphetamine and amphetamine (Limbril) (Adderall) Amoxapine (Asendin) Diazepam (Valium; Diastat) Aripiprazole (Abilify) Diethylpropion (No Brand Name) Armodafinil (Nuvigil) Dimenhydrinate (Dramamine) Asenapine (Saphris) Donepezil (Aricept) Aspirin/caffeine (BC Powder; Goody Doripenem (Doribax) Powder) Doxapram (Dopram) Atomoxetine (Strattera) Doxepin (Silenor) Baclofen (Gablofen; Lioresal) Droperidol (No Brand Name) Benzphetamine (Didrex; Regimex) Duloxetine (Cymbalta) Benztropine (Cogentin) Entacapone (Comtan) Bismuth Ergotamine and caffeine (Cafergot; subcitrate/metronidazole/tetracycline Migergot) (Pylera) Escitalopram (Lexapro) Bismuth subsalicylate (Pepto-Bismol) Fluoxetine (Prozac; Sarafem) -

Structural and Functional Neuroprotection in Glaucoma: Role of Galantamine-Mediated Activation of Muscarinic Acetylcholine Receptors
Citation: Cell Death and Disease (2010) 1, e27; doi:10.1038/cddis.2009.23 & 2010 Macmillan Publishers Limited All rights reserved 2041-4889/10 www.nature.com/cddis Structural and functional neuroprotection in glaucoma: role of galantamine-mediated activation of muscarinic acetylcholine receptors M Almasieh1, Y Zhou1, ME Kelly2, C Casanova3 and A Di Polo*,1 Glaucoma is the leading cause of irreversible blindness worldwide. Loss of vision due to glaucoma is caused by the selective death of retinal ganglion cells (RGCs). Treatments for glaucoma, limited to drugs or surgery to lower intraocular pressure (IOP), are insufficient. Therefore, a pressing medical need exists for more effective therapies to prevent vision loss in glaucoma patients. In this in vivo study, we demonstrate that systemic administration of galantamine, an acetylcholinesterase inhibitor, promotes protection of RGC soma and axons in a rat glaucoma model. Functional deficits caused by high IOP, assessed by recording visual evoked potentials from the superior colliculus, were improved by galantamine. These effects were not related to a reduction in IOP because galantamine did not change the pressure in glaucomatous eyes and it promoted neuronal survival after optic nerve axotomy, a pressure-independent model of RGC death. Importantly, we demonstrate that galantamine- induced ganglion cell survival occurred by activation of types M1 and M4 muscarinic acetylcholine receptors, while nicotinic receptors were not involved. These data provide the first evidence of the clinical potential -

Professor Jo Klaveness School of Pharmacy University of Oslo
SUPERVISORS Professor Jo Klaveness School of Pharmacy University of Oslo Associate professor Pål Rongved School of Pharmacy University of Oslo 1 TABLE OF CONTENTS TABLE OF CONTENTS 1 ABBREVIATIONS..........................................................................5 2 ABSTRACT.....................................................................................6 3 INTRODUCTION............................................................................7 3.1 Acetylcholine- a neurotransmitter Synthesis, release and inactivation ... 7 3.1.1 Structure of acetylcholinesterase .................................................................... 8 3.2 Anticholinesterases interfere with acetylcholine activity ........................ 9 3.3 Effects of anticholinesterases................................................................... 9 3.4 Different groups of anticholinesterases.................................................. 10 3.4.1 Short- acting anticholinesterases .................................................................. 10 3.4.2 Medium- duration anticholinesterases .......................................................... 11 3.4.3 Irreversible anticholinesterases..................................................................... 12 3.5 Nerve agents: Irreversible anticholinesterases....................................... 13 3.5.1 The dawn of a deadly weapon ...................................................................... 14 3.5.2 Biochemistry................................................................................................