Comparison of the Effects of DC031050, a Class III Antiarrhythmic Agent, on Herg Channel and Three Neuronal Potassium Channels
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
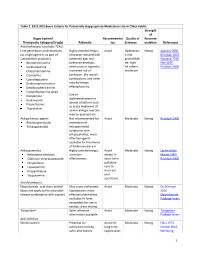
Table 2. 2012 AGS Beers Criteria for Potentially
Table 2. 2012 AGS Beers Criteria for Potentially Inappropriate Medication Use in Older Adults Strength of Organ System/ Recommendat Quality of Recomm Therapeutic Category/Drug(s) Rationale ion Evidence endation References Anticholinergics (excludes TCAs) First-generation antihistamines Highly anticholinergic; Avoid Hydroxyzin Strong Agostini 2001 (as single agent or as part of clearance reduced with e and Boustani 2007 combination products) advanced age, and promethazi Guaiana 2010 Brompheniramine tolerance develops ne: high; Han 2001 Carbinoxamine when used as hypnotic; All others: Rudolph 2008 Chlorpheniramine increased risk of moderate Clemastine confusion, dry mouth, Cyproheptadine constipation, and other Dexbrompheniramine anticholinergic Dexchlorpheniramine effects/toxicity. Diphenhydramine (oral) Doxylamine Use of diphenhydramine in Hydroxyzine special situations such Promethazine as acute treatment of Triprolidine severe allergic reaction may be appropriate. Antiparkinson agents Not recommended for Avoid Moderate Strong Rudolph 2008 Benztropine (oral) prevention of Trihexyphenidyl extrapyramidal symptoms with antipsychotics; more effective agents available for treatment of Parkinson disease. Antispasmodics Highly anticholinergic, Avoid Moderate Strong Lechevallier- Belladonna alkaloids uncertain except in Michel 2005 Clidinium-chlordiazepoxide effectiveness. short-term Rudolph 2008 Dicyclomine palliative Hyoscyamine care to Propantheline decrease Scopolamine oral secretions. Antithrombotics Dipyridamole, oral short-acting* May -

Dofetilide (Tikosyn): a New Drug to Control Atrial Fibrillation
CURRENT DRUG THERAPY WALID I. SALIBA, MD Section of Cardiac Electrophysiology and Pacing, CREDIT Department of Cardiology, Cleveland Clinic Dofetilide (Tikosyn): A new drug to control atrial fibrillation ABSTRACT OFETILIDE (Tikosyn), a new antiarrhyth- mic drug, can convert atrial fibrillation Dofetilide, a new class III antiarrhythmic agent, selectively and atrial flutter to sinus rhythm in approxi- blocks a specific cardiac potassium channel, lKr, increasing mately 30% of cases and maintain sinus the effective refractory period of the myocyte and thereby rhythm after electrical or pharmacologic con- terminating reentrant arrhythmias. Given orally, it appears version for up to 1 year in 60% to 70% of to effectively convert atrial fibrillation and atrial flutter to cases, without increasing the risk of sudden sinus rhythm and maintain sinus rhythm after conversion in death in patients at high risk. appropriately selected patients. This paper reviews the Such new drugs are needed, as many of the pharmacology of dofetilide, the evidence of its antiarrhythmic drugs in use up to now have effectiveness, and the appropriate precautions in using it. actually produced higher mortality rates in clinical trials than did placebo, or cause unac- ceptable side effects. KEY POINTS This article reviews the mechanism of action, safety, effectiveness, and clinical use of Dofetilide is generally well tolerated but like other class III dofetilide. drugs can cause torsades de pointes. The risk is dose- dependent and can be minimized by adjusting the dosage • PROBLEMS WITH PREVIOUS DRUGS according to creatinine clearance and QT interval, by excluding patients with known risk factors for long QT A variety of drugs have been used to terminate syndrome and torsades de pointes, and by starting or prevent atrial and ventricular arrhythmias, treatment in an inpatient monitored setting for the first 3 but their safety, efficacy, and tolerability in days. -

Drug Class Review Beta Adrenergic Blockers
Drug Class Review Beta Adrenergic Blockers Final Report Update 4 July 2009 Update 3: September 2007 Update 2: May 2005 Update 1: September 2004 Original Report: September 2003 The literature on this topic is scanned periodically. The purpose of this report is to make available information regarding the comparative effectiveness and safety profiles of different drugs within pharmaceutical classes. Reports are not usage guidelines, nor should they be read as an endorsement of, or recommendation for, any particular drug, use, or approach. Oregon Health & Science University does not recommend or endorse any guideline or recommendation developed by users of these reports. Mark Helfand, MD, MPH Kim Peterson, MS Vivian Christensen, PhD Tracy Dana, MLS Sujata Thakurta, MPA:HA Drug Effectiveness Review Project Marian McDonagh, PharmD, Principal Investigator Oregon Evidence-based Practice Center Mark Helfand, MD, MPH, Director Oregon Health & Science University Copyright © 2009 by Oregon Health & Science University Portland, Oregon 97239. All rights reserved. Final Report Update 4 Drug Effectiveness Review Project TABLE OF CONTENTS INTRODUCTION .......................................................................................................................... 6 Purpose and Limitations of Evidence Reports........................................................................................ 8 Scope and Key Questions .................................................................................................................... 10 METHODS................................................................................................................................. -

Arrhythmias in the Intensive Care Patient Hans-Joachim Trappe, Bodo Brandts and Peter Weismueller
Arrhythmias in the intensive care patient Hans-Joachim Trappe, Bodo Brandts and Peter Weismueller Purpose of review ventricular fibrillation. The use of automatic external Atrial fibrillation, atrial flutter, AV–nodal reentry tachycardia defibrillators by basic life support ambulance providers or first with rapid ventricular response, atrial ectopic tachycardia, and responder in early defibrillation programs has been associated preexcitation syndromes combined with atrial fibrillation or with a significant increase in survival rates. Drugs such as ventricular tachyarrhythmias are typical arrhythmias in intensive lidocaine, procainamide, sotalol, amiodarone, or magnesium care patients. Most frequently, the diagnosis of the underlying were recommended for treatment of ventricular arrhythmia is possible from the physical examination, the tachyarrhythmias in intensive care patients. Amiodarone is a response to maneuvers or drugs, and the 12-lead surface highly efficacious antiarrhythmic agent for many cardiac electrocardiogram. In all patients with unstable hemodynamics, arrhythmias, ranging from atrial fibrillation to malignant immediate DC-cardioversion is indicated. Conversion of atrial ventricular tachyarrhythmias, and seems to be superior to other fibrillation to sinus rhythm is possible using antiarrhythmic antiarrhythmic agents. drugs. Amiodarone has a conversion rate in atrial fibrillation of up to 80%. However, caution in the use of short-term Keywords administration of intravenous amiodarone in critically ill patients atrial fibrillation, -

Download Article (PDF)
Open Life Sci. 2018; 13: 335–339 Mini-Review Zhe An, Guang Yang, Xuanxuan Liu, Zhongfan Zhang, Guohui Liu* New progress in understanding the cellular mechanisms of anti-arrhythmic drugs https://doi.org/10.1515/biol-2018-0041 arrhythmia still require drugs to terminate the episode; Received May 4, 2018; accepted June 8, 2018 some symptomatic supraventricular and ventricular Abstract: Antiarrhythmic drugs are widely used, however, premature beats need to be controlled with drugs and to their efficacy is moderate and they can have serious prevent recurrence. In addition, some patients cannot be side effects. Even if catheter ablation is effective for the placed on ICDs or have radiofrequency ablation performed treatment of atrial fibrillation and ventricular tachycardia, due to economic limitation. To enable clinicians and antiarrhythmic drugs are still important tools for the pharmacists to rationally evaluate antiarrhythmic drugs, treatment of arrhythmia. Despite efforts, the development various antiarrhythmic drugs are briefly discussed. of antiarrhythmic drugs is still slow due to the limited Furthermore, we reviewed emerging antiarrhythmic drugs understanding of the role of various ionic currents. This currently undergoing clinical investigation or already review summarizes the new targets and mechanisms of approved for clinical use. antiarrhythmic drugs. Keywords: Antiarrhythmic drugs; New targets; 2 Classification of anti-arrhythmic Mechanism drugs According to the electrophysiology and mechanism of action of Purkinje fiber in vitro, antiarrhythmic drugs can 1 Introduction generally be divided into four categories: Class I sodium channel blockers, including three subclasses of A, B, Arrhythmia is a common and dangerous cardiovascular C. Type IA is a modest blockade of sodium channels, disease [1]. -

Optum Essential Health Benefits Enhanced Formulary PDL January
PENICILLINS ketorolac tromethamineQL GENERIC mefenamic acid amoxicillin/clavulanate potassium nabumetone amoxicillin/clavulanate potassium ER naproxen January 2016 ampicillin naproxen sodium ampicillin sodium naproxen sodium CR ESSENTIAL HEALTH BENEFITS ampicillin-sulbactam naproxen sodium ER ENHANCED PREFERRED DRUG LIST nafcillin sodium naproxen DR The Optum Preferred Drug List is a guide identifying oxacillin sodium oxaprozin preferred brand-name medicines within select penicillin G potassium piroxicam therapeutic categories. The Preferred Drug List may piperacillin sodium/ tazobactam sulindac not include all drugs covered by your prescription sodium tolmetin sodium drug benefit. Generic medicines are available within many of the therapeutic categories listed, in addition piperacillin sodium/tazobactam Fenoprofen Calcium sodium to categories not listed, and should be considered Meclofenamate Sodium piperacillin/tazobactam as the first line of prescribing. Tolmetin Sodium Amoxicillin/Clavulanate Potassium LOW COST GENERIC PREFERRED For benefit coverage or restrictions please check indomethacin your benefit plan document(s). This listing is revised Augmentin meloxicam periodically as new drugs and new prescribing LOW COST GENERIC naproxen kit information becomes available. It is recommended amoxicillin that you bring this list of medications when you or a dicloxacillin sodium CARDIOVASCULAR covered family member sees a physician or other penicillin v potassium ACE-INHIBITORS healthcare provider. GENERIC QUINOLONES captopril ANTI-INFECTIVES -

ANTIARRHYTHMIC DRUGS Geoffrey W
Chapter 24 ANTIARRHYTHMIC DRUGS Geoffrey W. Abbott and Roberto Levi HISTORICAL PERSPECTIVE BASIC PHARMACOLOGY Singh-Vaughan Williams Classification of Antiarrhythmic Drugs HISTORICAL PERSPECTIVE Sodium Channels and Class I Antiarrhythmic Drugs β Receptors and Class II Antiarrhythmics Potassium Channels and Class III Antiarrhythmic Drugs The heart, and more specifically the heartbeat, has through- Calcium Channels and Class IV Antiarrhythmics out history served as an indicator of well-being and disease, CLINICAL PHARMACOLOGY both to the physician and to the patient. Through one’s own Categories of Arrhythmogenic Mechanisms heartbeat, one can feel the physiologic manifestations of joy, CLINICAL APPLICATION thrills, fear, and passion; the rigors of a sprint or long- Class I—Sodium Channel Blockers distance run; the instantaneous effects of medications, recre- Class II—β Blockers ational drugs, or toxins; the adrenaline of a rollercoaster ride Class III—Potassium Channel Blockers or a penalty shootout in a World Cup final. Although the Class IV—Calcium Channel Blockers complexities of the heart continue to humble the scientists EMERGING DEVELOPMENTS and physicians who study it, the heart is unique in that, Molecular Genetics of Arrhythmias despite the complexity of its physiology and the richness of hERG Drug Interactions both visceral and romantic imagery associated with it, its Gene Therapy Guided by Molecular Genetics of Inherited function can be distilled down to that of a simple pump, the Arrhythmias function and dysfunction of which -

Management of Chronic Problems
MANAGEMENT OF CHRONIC PROBLEMS INTERACTIONS BETWEEN ALCOHOL AND DRUGS A. Leary,* T. MacDonald† SUMMARY concerned. Alcohol may alter the effects of the drug; drug In western society alcohol consumption is common as is may change the effects of alcohol; or both may occur. the use of therapeutic drugs. It is not surprising therefore The interaction between alcohol and drug may be that concomitant use of these should occur frequently. The pharmacokinetic, with altered absorption, metabolism or consequences of this combination vary with the dose of elimination of the drug, alcohol or both.2 Alcohol may drug, the amount of alcohol taken, the mode of affect drug pharmacokinetics by altering gastric emptying administration and the pharmacological effects of the drug or liver metabolism. Drugs may affect alcohol kinetics by concerned. Interactions may be pharmacokinetic or altering gastric emptying or inhibiting gastric alcohol pharmacodynamic, and while coincidental use of alcohol dehydrogenase (ADH).3 This may lead to altered tissue may affect the metabolism or action of a drug, a drug may concentrations of one or both agents, with resultant toxicity. equally affect the metabolism or action of alcohol. Alcohol- The results of concomitant use may also be principally drug interactions may differ with acute and chronic alcohol pharmacodynamic, with combined alcohol and drug effects ingestion, particularly where toxicity is due to a metabolite occurring at the receptor level without important changes rather than the parent drug. There is both inter- and intra- in plasma concentration of either. Some interactions have individual variation in the response to concomitant drug both kinetic and dynamic components and, where this is and alcohol use. -

Safety and Efficacy of Ibutilide in Cardioversion of Atrial Flutter And
J Am Board Fam Med: first published as 10.3122/jabfm.2011.01.080096 on 5 January 2011. Downloaded from CLINICAL REVIEW Safety and Efficacy of Ibutilide in Cardioversion of Atrial Flutter and Fibrillation Madhuri Nair, MD, Lekha K. George, MD, and Santhosh K. G. Koshy, MD This article reviews the safety and efficacy of ibutilide for use in patients with atrial fibrillation and flut- ter. Ibutilide, a class III antiarrhythmic agent, is primarily used for conversion of atrial flutter and fi- brillation and is a good alternative to electrical cardioversion. Ibutilide has a conversion rate of up to 75% to 80% in recent-onset atrial fibrillation and flutter; the conversion rate is higher for atrial flutter than for atrial fibrillation. It is also safe in the conversion of chronic atrial fibrillation/flutter among patients receiving oral amiodarone therapy. Ibutilide pretreatment facilitates transthoracic defibrilla- tion and decreases the energy requirement of electrical cardioversion by both monophasic and biphasic shocks. Pretreatment with ibutilide before electrical defibrillation has a conversion rate of 100% com- pared with 72% with no pretreatment. Ibutilide is also safe and efficient in the treatment of atrial fibril- lation in patients who have had cardiac surgery, and in accessory pathway–mediated atrial fibrillation where the conversion rate of ibutilide is as high as 95%. There is up to a 4% risk of torsade de pointes and a 4.9% risk of monomorphic ventricular tachycardia. Hence, close monitoring in an intensive care unit setting is warranted during and at least for 4 hours after drug infusion. The anticoagulation strat- egy is the same as for any other mode of cardioversion.(J Am Board Fam Med 2011;24:86–92.) Keywords: Antiarrhythmics, Arrhythmia, Atrial Fibrillation, Cardiovascular Disorders, Cardioversion, Drug Ther- copyright. -

Supplementary Materials
Supplementary Materials Table S1. The significant drug pairs in potential DDIs examined by the two databases. Micromedex Drugs.com List of drugs paired PK-PD Mechanism details 1. Amiodarone— PD Additive QT-interval prolongation Dronedarone 2. Amiodarone— PK CYP3A inhibition by Ketoconazole Ketoconazole 3. Ciprofloxacin— PD Additive QT-interval prolongation Dronedarone 4. Cyclosporine— PK CYP3A inhibition by Cyclosporine Dronedarone 5. Dronedarone— PK CYP3A inhibition by Erythromycin Erythromycin 6. Dronedarone— PD Additive QT-interval prolongation Flecainide 7. Dronedarone— PK CYP3A4 inhibition by Itraconazole Itraconazole 8. Dronedarone— PK Contraindication Major CYP3A inhibition by Ketoconazole Ketoconazole 9. Dronedarone— PD Additive QT-interval prolongation Procainamide PD 10. Dronedarone—Sotalol Additive QT-interval prolongation 11. Felodipine— PK CYP3A inhibition by Itraconazole Itraconazole 12. Felodipine— PK CYP3A inhibition by Ketoconazole Ketoconazole 13. Itraconazole— PK CYP3A inhibition by Itraconazole Nisoldipine 14. Ketoconazole— PK CYP3A inhibition by Ketoconazole Nisoldipine 15. Praziquantel— PK CYP induction by Rifampin Rifampin PD 1. Amikacin—Furosemide Additive or synergistic toxicity 2. Aminophylline— Decreased clearance of PK Ciprofloxacin Theophylline by Ciprofloxacin 3. Aminophylline— PK Decreased hepatic metabolism Mexiletine 4. Amiodarone— PD Additive effects on QT interval Ciprofloxacin 5. Amiodarone—Digoxin PK P-glycoprotein inhibition by Amiodarone 6. Amiodarone— PD, PK Major Major Additive effects on QT Erythromycin prolongation, CYP3A inhibition by Erythromycin 7. Amiodarone— PD, PK Flecainide Antiarrhythmic inhibition by Amiodarone, CYP2D inhibition by Amiodarone 8. Amiodarone— PK CYP3A inhibition by Itraconazole Itraconazole 9. Amiodarone— PD Antiarrhythmic inhibition by Procainamide Amiodarone 10. Amiodarone— PK CYP induction by Rifampin Rifampin PD Additive effects on refractory 11. Amiodarone—Sotalol potential 12. Amiodarone— PK CYP3A inhibition by Verapamil Verapamil 13. -

Efficacy and Safety of Ibutilide for the Cardioversion of Atrial Fibrillation: a Systematic Review and Meta- Analysis
Open Access Austin Surgery Case Reports Review Article Efficacy and Safety of Ibutilide for the Cardioversion of Atrial Fibrillation: A Systematic Review and Meta- Analysis Gong CC1, Tang Y2, Huang Y2 and Liu X2* 1Zhejiang University School of Medicine Children’s Abstract Hospital, China Background: Ibutilide has been approved for cardioversion of Atrial 2Department of Critical Care Medicine, the Affiliated Fibrillation (AF), but its side-effects include a high risk of torsade de pointes, Hospital of Guizhou Medical University, China besides, one recent meta-analysis showed ibutilide was inferior to vernakalant *Corresponding author: Liu X, Department of Critical for conversion (AF<7 days). Hence, the aim of this study is to evaluate the Care Medicine, the Affiliated Hospital of Guizhou Medical efficacy and safety of ibutilide for the cardioversion of AF within 90 days. University, China Methods: The Embase, PubMed, Web of Science, Cochrane Central Received: February 16, 2021; Accepted: March 17, databases and clinical trials.gov were comprehensively searched for relevant 2021; Published: March 24, 2021 studies from January 1991 to May 2020 using the keywords “ibutilide” and “atrial fibrillation”. Only Randomized Controlled Trials (RCTs) comparing ibutilide with placebo or other Anti-Arrhythmic Drugs (AADs) for the termination of AF (duration of AF ≤90 days) were included. The primary outcome was successful cardioversion in response to ibutilide versus placebo or other AADs within 4h. Related adverse events were defined as secondary outcomes. Results: A total of 1712 patients in 13 RCTs met the eligibility criteria. Four trials compared ibutilide to placebo; nine trials compared ibutilide to other active drugs. -

Australian Adverse Drug Reactions Bulletin, Volume 24 No 6
Prepared by the Adverse Drug Reactions Advisory Committee (ADRAC). Members of ADRAC are Associate Professor Duncan Topliss (Chair), Dr Vicki Kotsirilos, Professor David Isaacs, Dr Cecilie Lander, Professor John McNeil, Associate Professor Peter Pillans, Dr Simone Strasser, Dr Dana Wainwright AUSTRALIAN ADVERSE DRUG REACTIONS BULLETIN Volume 24, Number 6, December 2005 ! Medicines and QT prolongation ! Skin reactions with glucosamine ! Warfarin-induced skin necrosis Please report all suspected reactions to these Drugs of Current Interest Aripiprazole (Abilify) Levetiracetam (Keppra) Atomoxetine (Strattera) Pimecrolimus (Elidel) Ezetimibe (Ezetrol) Pregabalin (Lyrica) Fenofibrate (Lipidil) Reboxetine (Edronax) Iron sucrose (Venofer) Teriparatide (Fortéo) 1. Medicines and QT prolongation In the past decade, a number of medicines have Other, non-drug factors which may be associated been either withdrawn from the market or had their with QT prolongation include female sex, use restricted because of QT interval advanced age, bradycardia, cardiac failure, cardiac prolongation.1 The QT interval is considered to be ischaemia and electrolyte disturbances. prolonged if it is more than 450 msec after adjusting for heart rate (‘corrected QT interval’). Drug interactions are an important cause of QT prolongation and torsade de pointes, even in Drug-induced QT prolongation may be caused by healthy people with no risk factors. These blockade of cardiac potassium channels, and can interactions are of two types. The first involves the lead to a life-threatening polymorphic ventricular combined use of two or more drugs, each with a tachycardia known as torsade de pointes. As of QT prolonging effect, i.e. the simultaneous use of June 2005, ADRAC had received 140 reports of two drugs from the Table.