In the British Species of Papaver and Their Relation to Breeding Mechanisms and Ecology S
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Phylogeography of a Tertiary Relict Plant, Meconopsis Cambrica (Papaveraceae), Implies the Existence of Northern Refugia for a Temperate Herb
Article (refereed) - postprint Valtueña, Francisco J.; Preston, Chris D.; Kadereit, Joachim W. 2012 Phylogeography of a Tertiary relict plant, Meconopsis cambrica (Papaveraceae), implies the existence of northern refugia for a temperate herb. Molecular Ecology, 21 (6). 1423-1437. 10.1111/j.1365- 294X.2012.05473.x Copyright © 2012 Blackwell Publishing Ltd. This version available http://nora.nerc.ac.uk/17105/ NERC has developed NORA to enable users to access research outputs wholly or partially funded by NERC. Copyright and other rights for material on this site are retained by the rights owners. Users should read the terms and conditions of use of this material at http://nora.nerc.ac.uk/policies.html#access This document is the author’s final manuscript version of the journal article, incorporating any revisions agreed during the peer review process. Some differences between this and the publisher’s version remain. You are advised to consult the publisher’s version if you wish to cite from this article. The definitive version is available at http://onlinelibrary.wiley.com Contact CEH NORA team at [email protected] The NERC and CEH trademarks and logos (‘the Trademarks’) are registered trademarks of NERC in the UK and other countries, and may not be used without the prior written consent of the Trademark owner. 1 Phylogeography of a Tertiary relict plant, Meconopsis cambrica 2 (Papaveraceae), implies the existence of northern refugia for a 3 temperate herb 4 Francisco J. Valtueña*†, Chris D. Preston‡ and Joachim W. Kadereit† 5 *Área de Botánica, Facultad deCiencias, Universidad de Extremadura, Avda. de Elvas, s.n. -

The Alien Vascular Flora of Tuscany (Italy)
Quad. Mus. St. Nat. Livorno, 26: 43-78 (2015-2016) 43 The alien vascular fora of Tuscany (Italy): update and analysis VaLerio LaZZeri1 SUMMARY. Here it is provided the updated checklist of the alien vascular fora of Tuscany. Together with those taxa that are considered alien to the Tuscan vascular fora amounting to 510 units, also locally alien taxa and doubtfully aliens are reported in three additional checklists. The analysis of invasiveness shows that 241 taxa are casual, 219 naturalized and 50 invasive. Moreover, 13 taxa are new for the vascular fora of Tuscany, of which one is also new for the Euromediterranean area and two are new for the Mediterranean basin. Keywords: Vascular plants, Xenophytes, New records, Invasive species, Mediterranean. RIASSUNTO. Si fornisce la checklist aggiornata della fora vascolare aliena della regione Toscana. Insieme alla lista dei taxa che si considerano alieni per la Toscana che ammontano a 510 unità, si segnalano in tre ulteriori liste anche i taxa che si ritengono essere presenti nell’area di studio anche con popolazioni non autoctone o per i quali sussistono dubbi sull’effettiva autoctonicità. L’analisi dello status di invasività mostra che 241 taxa sono casuali, 219 naturalizzati e 50 invasivi. Inoltre, 13 taxa rappresentano una novità per la fora vascolare di Toscana, dei quali uno è nuovo anche per l’area Euromediterranea e altri due sono nuovi per il bacino del Mediterraneo. Parole chiave: Piante vascolari, Xenofte, Nuovi ritrovamenti, Specie invasive, Mediterraneo. Introduction establishment of long-lasting economic exchan- ges between close or distant countries. As a result The Mediterranean basin is considered as one of this context, non-native plant species have of the world most biodiverse areas, especially become an important component of the various as far as its vascular fora is concerned. -

I1.3 Arable Land with Unmixed Crops Grown by Low-Intensity Agricultural Methods
European Red List of Habitats - Screes Habitat Group I1.3 Arable land with unmixed crops grown by low-intensity agricultural methods Summary Species-rich arable weed vegetation, characterised by now rare native and archaeophyte annual plants, is a survivor of ancient low-intensity agriculture and widely distributed across Europe but survives in abundance only in the mountainous areas of the Mediterranean region, particularly Italy. Since the intensification of agricultural practices in the middle of the 20th century, much of it subsidised by agri- environment funding, a significant decrease both in quantity and quality has been observed caused by excessive use of herbicides, biocides, and chemical fertilizers, improved seed-cleaning methods, sowing highly productive and competitive varieties of cereals, and removal of refugial habitats in the landscape due to merging of small fields into large ones. Such threats continue in many places, especially threatening surviving outliers. A large scale improvement of quantity and quality may require a revision of Common Agricultural Policy and agri-environment funding schemes and promotion of restoration initiatives. Synthesis Despite variable data quality, a lack of data from several countries and different interpretations of the habitat definition, the decreases in quantity and quality have been calculated using the territorial data from a sufficient number of countries to build an overall European average. Due to a large decrease in area over the last 50 years, the habitat qualifies for category Endangered under criterion A1. All countries except Italy (-20% to -40%) and Switzerland (-37.5%) reported a decrease in area between -50% and -99%. If the Italian and Swiss data were neglected, the overall assessment would result in category Critically Endangered. -

Characteristic of Pseudomonas Syringae Pv. Atrofaciens Isolated from Weeds of Wheat Field
applied sciences Article Characteristic of Pseudomonas syringae pv. atrofaciens Isolated from Weeds of Wheat Field Liudmyla Butsenko 1, Lidiia Pasichnyk 1 , Yuliia Kolomiiets 2, Antonina Kalinichenko 3,* , Dariusz Suszanowicz 3 , Monika Sporek 4 and Volodymyr Patyka 1 1 Zabolotny Institute of Microbiology and Virology, National Academy of Sciences of Ukraine, 03143 Kyiv, Ukraine; [email protected] (L.B.); [email protected] (L.P.); [email protected] (V.P.) 2 Department of Ecobiotechnology and Biodiversity, National University of Life and Environmental Sciences of Ukraine, 03143 Kyiv, Ukraine; [email protected] 3 Institute of Environmental Engineering and Biotechnology, University of Opole, 45-040 Opole, Poland; [email protected] 4 Institute of Biology, University of Opole, 45-040 Opole, Poland; [email protected] * Correspondence: [email protected]; Tel.: +48-787-321-587 Abstract: The aim of this study was the identification of the causative agent of the basal glume rot of wheat Pseudomonas syringae pv. atrofaciens from the affected weeds in wheat crops, and determination of its virulent properties. Isolation of P. syringae pv. atrofaciens from weeds of wheat crops was carried out by classical microbiological methods. To identify isolated bacteria, their morphological, cultural, biochemical, and serological properties as well as fatty acids and Random Amplification of Polymorphic DNA (RAPD)-PCR (Polymerase chain reaction) profiles with the OPA-13 primer were studied. Pathogenic properties were investigated by artificial inoculation of wheat plants and weed plants, from which bacteria were isolated. For the first time, bacteria that are virulent both for weeds and wheat were isolated from weeds growing in wheat crops. -

Seed Dormancy and Germination in an Ecological Context ANNUAL WEEDS Comparative Studies of Annual Weeds
Linköping Studies in Science and Technology SEED DORMANCY AND Dissertation No. 1088 GERMINATION IN AN ECOLOGICAL CONTEXT COMPARATIVE STUDIES OF Seed dormancy and germination in an ecological context ANNUAL WEEDS comparative studies of annual weeds Laila M. Karlsson LAILA M. KARLSSON Department of Physics, Chemistry and Biology Division of Ecology Linköping University SE-581 83 Linköping, Sweden Contents List of papers iv Abstract v Frövila och groning i ett ekologiskt sammanhang — jämförande studier av annuella ogräs: sammanfattning (in Swedish) vi Overview: Seed dormancy and germination in an ecological context — comparative studies of annual weeds 1 1. Introduction 1 1.1. Weeds and germination ecology 1 1.2. Germination 2 1.3. Germination timing 6 1.4. Seed dormancy 9 1.5. Comparisons 13 1.6. Aim 14 2. Methods 14 3. Results 16 3.1. General 16 3.2. Papaver 16 3.3. Lamium 18 3.4. Asteraceae 19 4. Discussion 19 4.1. Experimental methods 19 4.2. Variations within species 20 4.3. Describing and comparing dormancy and germination 22 4.3.1. Purpose 22 4.3.2. Statistics 23 4.3.3. Seed dormancy description 25 4.3.4. Picturing general responses 29 4.4. Comparisons of seed dormancy and germination 31 4.4.1. Dormancy pattern 31 4.4.2. Germination preferences 33 4.4.3. Dormancy strength 34 Seed dormancy and germination in an ecological context 4.5. Implications for the field 35 – comparative studies of annual weeds 4.5.1. Germination studies vs emergence in nature 35 4.5.2. Papaver 36 4.5.3. -

Mohn (Papaver Somniferum
Mohn (Papaver somniferum L.) Synonyme (Namen dieser Pflanze in allen 76 Sprachen anzeigen) pharmazeutisch Semen Papaveris Albanisch Lulëkuqe Μήκων Altgriechisch Mekon ፓፓ Amharisch Papi , Arabisch , Khashkhash, Abu an-num, Abu al-num, Abu an-noom, Abu al-noom Մեկոն, Մեկոնի Կուտ Armenisch Mekon, Megon; Mekoni Kut, Megoni Good (Samen) Assamesisch Xaş-xaş Azeri Хаш-хаш Baskisch Lobelarr Bengali Градински мак, Опиев мак, Маково семе Bulgarisch Gradinski mak, Opiev mak; Makovo seme (Samen) Burmesisch Chinesisch 櫻粟殼 [yìng suhk hohk] (Kantonesisch) Ying suhk hohk Chinesisch 櫻粟殼 [yīng sù qiào], 罂粟 [yīng sù] (Mandarin) Ying su qiao, Ying su Dänisch Opiumvalmue (Pflanze); Birkes, Valmue-frø (Samen) Deutsch Schlafmohn, Gartenmohn, Ölmohn, Opiummohn, , Dhivehi Afihun, Kaskasaa Englisch Poppy, Opium poppy, Garden poppy Esperanto Papavo, Papavosemo Estnisch Magun, Unimagun, Moon Farsi Khash-khash, Shagheyegh Finnisch Unikko, Oopiumiunikko Französisch Pavot des jardins, Pavot somnifère, Pavot à opium Gälisch Codalion, Paipin Galizisch Mapoula, Sementes de Mapoula, Adormideira, Durmideira ყაყაჩო, ყაყაჩოს თესლი, ხოშხოში Georgisch Khoshkhoshi, Q’aq’acho, Xoshxoshi, Qaqacho; Q’aq’achos tesli, Qaqachos tesli (Samen) Παπαρούνα, Αθιόνι Griechisch Paparouna, Afioni Gujarati Hebräisch Pereg Hindi Irisch Poipín Isländisch Valmúafræ, Birki Italienisch Papavero, Papavero sonnifero 芥子, 罌粟 けし Japanisch ケシ, ポピー Keshi, Papi , Jiddisch Mondl, Mon Kannada Көкнәр Kasachisch Köknär Kashmiri Khash-khash Katalanisch Cascall, Herba dormidora 아편, 포피, 양귀비 Koreanisch Apyeon, Apyon, -

44 * Papaveraceae 1
44 * PAPAVERACEAE 1 Dennis I Morris 2 Annual or perennial herbs, rarely shrubs, with latex generally present in tubes or sacs throughout the plants. Leaves alternate, exstipulate, entire or more often deeply lobed. Flowers often showy, solitary at the ends of the main and lateral branches, bisexual, actinomorphic, receptacle hypogynous or perigynous. Sepals 2–3(4), free or joined, caducous. Petals (0–)4–6(–12), free, imbricate and often crumpled in the bud. Stamens usually numerous, whorled. Carpels 2-many, joined, usually unilocular, with parietal placentae which project towards the centre and sometimes divide the ovary into several chambers, ovules numerous. Fruit usually a capsule opening by valves or pores. Seeds small with crested or small raphe or with aril, with endosperm. A family of about 25 genera and 200 species; cosmopolitan with the majority of species found in the temperate and subtropical regions of the northern hemisphere. 6 genera and 15 species naturalized in Australia; 4 genera and 9 species in Tasmania. Papaveraceae are placed in the Ranunculales. Fumariaceae (mostly temperate N Hemisphere, S Africa) and Pteridophyllaceae (Japan) are included in Papaveraceae by some authors: here they are retained as separate families (see Walsh & Norton 2007; Stevens 2007; & references cited therein). Synonymy: Eschscholziaceae. Key reference: Kiger (2007). External resources: accepted names with synonymy & distribution in Australia (APC); author & publication abbre- viations (IPNI); mapping (AVH, NVA); nomenclature (APNI, IPNI). 1. Fruit a globular or oblong capsule opening by pores just below the stigmas 2 1: Fruit a linear capsule opening lengthwise by valves 3 2. Stigmas joined to form a disk at the top of the ovary; style absent 1 Papaver 2: Stigmas on spreading branches borne on a short style 2 Argemone 3. -

The Island of Cyprus
The Island of Cyprus Naturetrek Tour Report 29 March - 5th April 2014 Report compiled by Cliff Waller Naturetrek Cheriton Mill Cheriton Alresford Hampshire SO24 0NG England T: +44 (0)1962 733051 F: +44 (0)1962 736426 E: [email protected] W: www.naturetrek.co.uk Tour Report The Island of Cyprus Tour Leader: Yiannis Christofides Local Leader & Botanist Cliff Waller Naturetrek Ornithologist Participants: Kath Edwards Malcolm Forbes Pauline Grimshaw Hilary Homer Ian Homer Karen Mayer Annette Langlois Chris Meek Dave Meek Joanna O’Brien Paul O’Brien Bob Snellgrove Sandra Snellgrove Judith Stiffin Jill Wilson Day 1 Saturday 29th March London to Larnaca Everyone had an early but comfortable four hour flight from Gatwick to Paphos. We arrived more or less on time at 1.30 pm and the customs and immigration formalities were brief, so we were soon able to be greeted by Yiannis, our local guide and botanist and see our first new common birds, Hooded Crow and Wood Pigeon. It was only a 20 minute drive to our comfortable hotel and after quickly settling in we set off at 3.15 pm to walk to the Tombs of the Kings, about three quarters of a mile away, seeing Iris Gynandriris sisyrinchium near the hotel and our first Sardinian Warbler along the way. On arriving at the tombs we found a strong onshore wind was keeping any birds very low and in cover, but we did see a number of Crested Lark and European Wheatear, as well as less common species such as Spanish Sparrow and Cretzschmar’s Bunting, while brief views were also obtained of Isabelline Wheatear. -
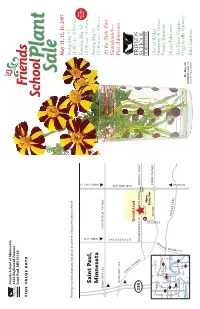
2007 Catalog
Friends School of Minnesota 1365 Englewood Avenue Saint Paul, MN 55104 TIME VALUE DATA May 11, 12, 13, 2007 Friday,May 11 If you have received a duplicate copy, please let us know, and pass the extra to a friend! 11:00 A.M.–8:00 P.M. New Saturday Saturday,May 12 Hours Saint Paul, 10:00 A.M.–6:00 P.M. Sunday,May 13 FROM 35W Minnesota FROM HWY 36 12:00 NOON–4:00 P.M. FROM HWY 280 LARPENTEUR AVENUE At the State Fair Grandstand— FROM HWY 280 Free Admission C O M O CLEVELAND AVE A SNELLING AVE V E Grandstand N U 280 E COMMONWEALTH DAN PATCH Main MIDWAY PKWY Gate P Minn. State Fair 94 Coliseum COMO AVENUE 35W White Shoreview Glacial Ridge Brooklyn Ctr Bear Lake 694 35E E U CANFIELD Growers: A Green Plymouth Crystal 94 Roseville N 36 E 494 Snelling Ave. 694 V 169 Saint Paul Family Business 280 A 394 35E 100 94 D Minnetonka Minneapolis E N N Woodbury ERGY Hosta Takeover! O P Edina 494 ARK 62 M Richfield Y 61 Eden 494 Prairie A Are These Veggies 35W Inver Grove R Heights Bloomington Eagan FROM 94 Organically Grown? 52 Mr. Majestic Shakopee 35E Burnsville marigold, page 12 Photo by Nancy Scherer Bird Gardens 18th Annual Friends School Plant Sale May 11, 12 and 13, 2007 Friday 11:00 A.M.–8:00 P.M.• Saturday 10:00 A.M.–6:00 P.M. Sunday 12:00 NOON–4:00 P.M.Sunday is half-price day at the Minnesota State Fair Grandstand Friends School of Minnesota Thank you for supporting Friends School of Minnesota by purchasing plants at our sale. -

Remarkable Changes of Weed Species in Spanish Cereal Fields from 1976 to 2007 Cirujeda, Aibar, Zaragoza
Remarkable changes of weed species in Spanish cereal fields from 1976 to 2007 Cirujeda, Aibar, Zaragoza To cite this version: Cirujeda, Aibar, Zaragoza. Remarkable changes of weed species in Spanish cereal fields from 1976 to 2007. Agronomy for Sustainable Development, Springer Verlag/EDP Sciences/INRA, 2011, 31 (4), pp.675-688. 10.1007/s13593-011-0030-4. hal-00930501 HAL Id: hal-00930501 https://hal.archives-ouvertes.fr/hal-00930501 Submitted on 1 Jan 2011 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Agronomy Sust. Developm. (2011) 31:675–688 DOI 10.1007/s13593-011-0030-4 RESEARCH ARTICLE Remarkable changes of weed species in Spanish cereal fields from 1976 to 2007 Alicia Cirujeda & Joaquín Aibar & Carlos Zaragoza Accepted: 4 January 2011 /Published online: 18 March 2011 # INRA and Springer Science+Business Media B.V. 2011 Abstract Management practices, geographical gradients Convolvulus arvensis found in more than half of the and climatic factors are factors explaining weed species surveyed fields. L. rigidum was related to dryland, while composition and richness in cereal fields from Northern and the other species were found overall. Furthermore, we Central Europe. -

Piedmont, North Italy)
Italian Botanist 5: 57–69Contribution (2018) to the floristic knowledge of the head of the Po Valley... 57 doi: 10.3897/italianbotanist.5.24546 RESEARCH ARTICLE http://italianbotanist.pensoft.net Contribution to the floristic knowledge of the head of the Po Valley (Piedmont, north Italy) Daniela Bouvet1, Annalaura Pistarino2, Adriano Soldano3, Enrico Banfi4, Massimo Barbo5, Fabrizio Bartolucci6,7, Maurizio Bovio8, Laura Cancellieri9, Fabio Conti6,7, Romeo Di Pietro10, Francesco Faraoni11, Simonetta Fascetti12, Gabriele Galasso4, Carmen Gangale13, Edda Lattanzi14, Simonetta Peccenini15, Enrico Vito Perrino16, Roberto Rizzieri Masin17, Vito Antonio Romano12, Leonardo Rosati13, Giovanni Salerno18, Adriano Stinca19,20, Agnese Tilia21, Dimitar Uzunov22 1 Department of Life Sciences and Systems Biology, University of Turin, Viale P.A. Mattioli 25, 10125 Turin, Italy 2 Regional Museum of Natural Sciences, Via G. Giolitti 36, 10123 Turin, Italy 3 Largo Brigata Cagliari 6, 13100 Vercelli, Italy 4 Museum of Natural History, Botanical Department, Corso Venezia 55, 20121 Mi- lan, Italy 5 Via V. Alfieri 10, 33100 Udine, Italy6 School of Biosciences and Veterinary Medicine, University of Camerino, Italy 7 Apennines Floristic Research Center, San Colombo, Via Prov.le Km 4,2, 67021 Barisciano (L’Aquila), Italy 8 Via Saint Martin de Corléans 151, 11100 Aosta, Italy 9 Department of Agriculture and Forestry Science, Tuscia University, Via San Camillo de Lellis, 01100 Viterbo, Italy 10 Department of Plan- ning, Design, and Technology of Architecture, Sapienza University of Rome, Via Flaminia 72, 00196 Rome, Italy 11 Via Rubattino 6, 00153 Rome, Italy 12 School of Agricultural, Forestry, Food and Environmental Sciences, Basilicata University, Via Ateneo Lucano 10, 85100 Potenza, Italy 13 Museum of Natural History and Botanic Garden, University of Calabria, Loc. -

The Effects of Secondary Metabolites of Rangeland and Pasture Plants on the Animal Health in Mediterranean Ecological Conditions
Journal of US-China Medical Science 16 (2019) 63-72 doi: 10.17265/1548-6648/2019.01.003 D DAVID PUBLISHING The Effects of Secondary Metabolites of Rangeland and Pasture Plants on the Animal Health in Mediterranean Ecological Conditions Emre Kara and Mustafa Sürmen Departmant of Field Crops, Faculty of Agriculture, Aydın Adnan Menderes University, Aydın 09100, Turkey Abstract: Grazed areas including rangelands and pastures cover approximately 51% of the Earth’s continental surface. The plant species which are in these areas which form a wide biodiversity with living and non-living organisims, stand out with both as a forage source and medical use. In this natural habitat, plants are surrounded by a large number of potential enemies. Since plants are not able to escape from these enemies (herbivores and pathogenes), plants produce substances which are not functional for growing and develop but protect plants against external factors as defence mechanisms. These substances are called secondary metabolites. These substances which are seen differently from the primer metabolites, possess charachteristics specific to each plant species. Secondary metabolites of plant origin can be divided into three main groups as terpens, phenolic compounds and nitrogenous compounds. Terpenes have negative effects on many mammals and insects, because they are toxic. The lignin, a phenolic compound, is located in the plant cell wall and has a particularly digestibility-lowering effect. In addition, flavanoids, which are again phenolic compounds, are thought to be produced against harmful effects of UV rays and pathogen invasion. Tannins, another phenolic compounds, are generally toxic. When plants which have tannins are grazed by herbivorous, they significantly reduce chances of growth and survival of animals.