A Survey of Insecticide Resistance-Conferring Mutations in Multiple Targets in Anopheles Sinensis Populations Across Sichuan, Ch
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Lithofacies Palaeogeography of the Late Permian Wujiaping Age in the Middle and Upper Yangtze Region, China
Journal of Palaeogeography 2014, 3(4): 384-409 DOI: 10.3724/SP.J.1261.2014.00063 Lithofacies palaeogeography and sedimentology Lithofacies palaeogeography of the Late Permian Wujiaping Age in the Middle and Upper Yangtze Region, China Jin-Xiong Luo*, You-Bin He, Rui Wang School of Geosciences, Yangtze University, Wuhan 430100, China Abstract The lithofacies palaeogeography of the Late Permian Wujiaping Age in Middle and Upper Yangtze Region was studied based on petrography and the “single factor analysis and multifactor comprehensive mapping” method. The Upper Permian Wujiaping Stage in the Middle and Upper Yangtze Region is mainly composed of carbonate rocks and clastic rocks, with lesser amounts of siliceous rocks, pyroclastic rocks, volcanic rocks and coal. The rocks can be divided into three types, including clastic rock, clastic rock-limestone and lime- stone-siliceous rock, and four fundamental ecological types and four fossil assemblages are recognized in the Wujiaping Stage. Based on a petrological and palaeoecological study, six single factors were selected, namely, thickness (m), content (%) of marine rocks, content (%) of shallow water carbonate rocks, content (%) of biograins with limemud, content (%) of thin- bedded siliceous rocks and content (%) of deep water sedimentary rocks. Six single factors maps of the Wujiaping Stage and one lithofacies palaeogeography map of the Wujiaping Age were composed. Palaeogeographic units from west to east include an eroded area, an alluvial plain, a clastic rock platform, a carbonate rock platform where biocrowds developed, a slope and a basin. In addition, a clastic rock platform exists in the southeast of the study area. Hydro- carbon source rock and reservoir conditions were preliminarily analyzed based on lithofacies palaeogeography. -

Sichuan Q I N G H a I G a N S U Christian Percentage of County/City Ruo'ergai
Sichuan Q i n g h a i G a n s u Christian Percentage of County/City Ruo'ergai Shiqu Jiuzhaigou S h a a n x i Hongyuan Aba Songpan Chaotian Qingchuan Nanjiang Seda Pingwu Lizhou Rangtang Wangcang Dege Heishui Zhaohua Tongjiang Ma'erkang Ganzi Beichuan Jiangyou Cangxi Wanyuan Mao Jiange Bazhou Enyang Zitong Pingchang Luhuo Jinchuan Li Anzhou Youxian Langzhong Xuanhan Mianzhu Yilong Shifang Fucheng Tongchuan Baiyu Luojiang Nanbu Pengzhou Yangting Xiaojin Jingyang Santai Yingshan Dachuan Danba Dujiangyan Xichong Xinlong Wenchuan Guanghan Peng'an Kaijiang Daofu Shehong Shunqing Qu Pi Xindu Zhongjiang Gaoping Chongzhou Jialing DayiWenjiang Jintang Pengxi Guang'an Dazhu Lushan Daying Yuechi Qianfeng Shuangliu Chuanshan Baoxing Qionglai Huaying T i b e t Batang Xinjin Jianyang Anju Wusheng Pujiang Kangding Pengshan Lezhi Linshui Mingshan Yanjiang Tianquan DanlengDongpo H u b e i Litang Yajiang Yucheng Renshou Anyue Yingjing Qingshen Zizhong Luding Jiajiang Jingyan Hongya Shizhong Weiyuan Dongxing Hanyuan Emeishan Rong Shizhong WutongqiaoGongjing Da'an Longchang C h o n g q i n g Xiangcheng Shimian Jinkouhe Shawan Ziliujing Yantan Ebian Qianwei Lu Jiulong Muchuan Fushun Daocheng Ganluo Longmatan Derong Xuzhou NanxiJiangyang Mabian Pingshan Cuiping Hejiang Percent Christian Naxi Mianning Yuexi Jiang'an Meigu Changning (County/City) Muli Leibo Gao Gong Xide Xingwen 0.8% - 3% Zhaojue Junlian Xuyong Gulin Chengdu area enlarged 3.1% - 4% Xichang Jinyang Qingbaijiang Yanyuan Butuo Pi Puge Xindu 4.1% - 5% Dechang Wenjiang Y u n n a n Jinniu Chenghua Qingyang 5.1% - 6% Yanbian Ningnan Miyi G u i z h o u Wuhou Longquanyi 6.1% - 8.8% Renhe Jinjiang Xi Dong Huidong Shuangliu Renhe Huili Disputed boundary with India Data from Asia Harvest, www.asiaharvest.org. -

Expressways, GDP, and the Environment: the Case of China
Expressways, GDP, and the Environment: The Case of China Guojun He Hong Kong University of Science and Technology [email protected] Yang Xie University of California, Riverside [email protected] Bing Zhang Nanjing University [email protected] We are indebted to the Editor in Chief, Andrew Foster, and the anonymous referee for their valuable suggestions. We thank Hunt Allcott, Michael Anderson, Richard Arnott, Chong-En Bai, Michael Bates, David Brady, Nathaniel Baum-Snow, Cyndi Berck, Peter Berck, Judd Boomhower, David Brady, Jimmy Chan, Gordon Dahl, Anil Deolalikar, Alain de Janvry, Ozkan Eren, Thibault Fally, Jingting Fan, Fred Finan, Shihe Fu, Joshua Graff Zivin, Michael Greenstone, Jie He, Steven Helfand, Sarojini Hirshleifer, Wei Huang, Ruixue Jia, Larry Karp, Bree Lang, Matt Lang, Bryan Leonard, Weijia Li, Jeremy Magruder, Aprajit Mahajan, John Matsusaka, Daniel McMillen, Helene Ollivier, Albert Park, Martino Pelli, Jeffrey Perloff, Obie Porteous, Han Qi, David Rapson, Elisabeth Sadoulet, Ruoyao Shi, Leo Simon, Michael Song, Kenneth Small, Qu Tang, Itai Trilnick, Reed Walker, Shaoda Wang, Brian Wright, Yanhui Wu, Yiqing Xu, Jia Yan, and participants in seminars and workshops at CU Boulder, CUHK, Fudan University, HKBU, HKUST, Peking University, Shanghai Jiao Tong University, Singapore Management University, Tsinghua University, UC Berkeley, UC Riverside, Université de Sherbrooke, and USC and the 2017 AERE Summer Conference, CAERE Annual Conference, CES Annual Conference, EAERE Annual Conference, Econometric Society Asian Meeting, and Fudan–UC Social Science and China Studies Young Scholar Conference for their valuable comments. Yuhang Pan, Xiaoxiao Shen, Chun Wai Cheung, Ziteng Lei, Tingjun Man, and Jing Yang offered excellent research assistance. -

The Survey on the Distribution of MC Fei and Xiao Initial Groups in Chinese Dialects
IALP 2020, Kuala Lumpur, Dec 4-6, 2020 The Survey on the Distribution of MC Fei and Xiao Initial Groups in Chinese Dialects Yan Li Xiaochuan Song School of Foreign Languages, School of Foreign Languages, Shaanxi Normal University, Shaanxi Normal University Xi’an, China /Henan Agricultural University e-mail: [email protected] Xi’an/Zhengzhou, China e-mail:[email protected] Abstract — MC Fei 非 and Xiao 晓 initial group discussed in this paper includes Fei 非, Fu groups are always mixed together in the southern 敷 and Feng 奉 initials, but does not include Wei part of China. It can be divided into four sections 微, while MC Xiao 晓 initial group includes according to the distribution: the northern area, the Xiao 晓 and Xia 匣 initials. The third and fourth southwestern area, the southern area, the class of Xiao 晓 initial group have almost southeastern area. The mixing is very simple in the palatalized as [ɕ] which doesn’t mix with Fei northern area, while in Sichuan it is the most initial group. This paper mainly discusses the first extensive and complex. The southern area only and the second class of Xiao and Xia initials. The includes Hunan and Guangxi where ethnic mixing of Fei and Xiao initials is a relatively minorities gather, and the mixing is very recent phonetic change, which has no direct complicated. Ancient languages are preserved in the inheritance with the phonological system of southeastern area where there are still bilabial Qieyun. The mixing mainly occurs in the southern sounds and initial consonant [h], but the mixing is part of the mainland of China. -

December 1998
JANUARY - DECEMBER 1998 SOURCE OF REPORT DATE PLACE NAME ALLEGED DS EX 2y OTHER INFORMATION CRIME Hubei Daily (?) 16/02/98 04/01/98 Xiangfan C Si Liyong (34 yrs) E 1 Sentenced to death by the Xiangfan City Hubei P Intermediate People’s Court for the embezzlement of 1,700,00 Yuan (US$20,481,9). Yunnan Police news 06/01/98 Chongqing M Zhang Weijin M 1 1 Sentenced by Chongqing No. 1 Intermediate 31/03/98 People’s Court. It was reported that Zhang Sichuan Legal News Weijin murdered his wife’s lover and one of 08/05/98 the lover’s relatives. Shenzhen Legal Daily 07/01/98 Taizhou C Zhang Yu (25 yrs, teacher) M 1 Zhang Yu was convicted of the murder of his 01/01/99 Zhejiang P girlfriend by the Taizhou City Intermediate People’s Court. It was reported that he had planned to kill both himself and his girlfriend but that the police had intervened before he could kill himself. Law Periodical 19/03/98 07/01/98 Harbin C Jing Anyi (52 yrs, retired F 1 He was reported to have defrauded some 2600 Liaoshen Evening News or 08/01/98 Heilongjiang P teacher) people out of 39 million Yuan 16/03/98 (US$4,698,795), in that he loaned money at Police Weekend News high rates of interest (20%-60% per annum). 09/07/98 Southern Daily 09/01/98 08/01/98 Puning C Shen Guangyu D, G 1 1 Convicted of the murder of three children - Guangdong P Lin Leshan (f) M 1 1 reported to have put rat poison in sugar and 8 unnamed Us 8 8 oatmeal and fed it to the three children of a man with whom she had a property dispute. -

3 PRESENT CONDITION of ROADS and TRANSPORT 3.1 Condition of Road Facilities 3.1.1 Existing Road Network and Classification of R
3 PRESENT CONDITION OF ROADS AND TRANSPORT 3.1 Condition of Road Facilities 3.1.1 Existing Road Network and Classification of Roads (1) Primary Road Network in the Study Area As shown in Figure 3.1.1, the existing Primary Road network in the Study Area considers the Tianfu Square as its center and is made up of ring roads and radial roads. There are 5 ring roads at present including the existing roads and roads under construction. The existing roads include the Inner Ring Road with a radius of 2 km, the 1st Ring Road with a radius of 3 km, and the 2nd Ring Road with a radius of 4 km. There are two roads under construction, namely the 3rd Ring Road with a radius of 7 km, and the Outer Ring Road with a radius of 11 km. The construction of these two roads is underway, and most exits for expressways and main carriageway have been constructed already. It is expected that the 3rd Ring Road and the Outer Ring Road will be completed by 2001 and 2002, respectively. The Inner Ring Road has four lanes for both directions with a total road width of 25 m. The 1st Ring Road and 2nd Ring Road have not only four lanes for both directions but also a segregated bicycle lane on both sides with a total width of 40 m. Furthermore, the 3rd Ring Road and the Outer Ring Road under construction have 8 lanes and 6 lanes for both directions, respectively, with a total width of 80m. -
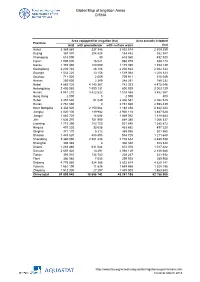
Global Map of Irrigation Areas CHINA
Global Map of Irrigation Areas CHINA Area equipped for irrigation (ha) Area actually irrigated Province total with groundwater with surface water (ha) Anhui 3 369 860 337 346 3 032 514 2 309 259 Beijing 367 870 204 428 163 442 352 387 Chongqing 618 090 30 618 060 432 520 Fujian 1 005 000 16 021 988 979 938 174 Gansu 1 355 480 180 090 1 175 390 1 153 139 Guangdong 2 230 740 28 106 2 202 634 2 042 344 Guangxi 1 532 220 13 156 1 519 064 1 208 323 Guizhou 711 920 2 009 709 911 515 049 Hainan 250 600 2 349 248 251 189 232 Hebei 4 885 720 4 143 367 742 353 4 475 046 Heilongjiang 2 400 060 1 599 131 800 929 2 003 129 Henan 4 941 210 3 422 622 1 518 588 3 862 567 Hong Kong 2 000 0 2 000 800 Hubei 2 457 630 51 049 2 406 581 2 082 525 Hunan 2 761 660 0 2 761 660 2 598 439 Inner Mongolia 3 332 520 2 150 064 1 182 456 2 842 223 Jiangsu 4 020 100 119 982 3 900 118 3 487 628 Jiangxi 1 883 720 14 688 1 869 032 1 818 684 Jilin 1 636 370 751 990 884 380 1 066 337 Liaoning 1 715 390 783 750 931 640 1 385 872 Ningxia 497 220 33 538 463 682 497 220 Qinghai 371 170 5 212 365 958 301 560 Shaanxi 1 443 620 488 895 954 725 1 211 648 Shandong 5 360 090 2 581 448 2 778 642 4 485 538 Shanghai 308 340 0 308 340 308 340 Shanxi 1 283 460 611 084 672 376 1 017 422 Sichuan 2 607 420 13 291 2 594 129 2 140 680 Tianjin 393 010 134 743 258 267 321 932 Tibet 306 980 7 055 299 925 289 908 Xinjiang 4 776 980 924 366 3 852 614 4 629 141 Yunnan 1 561 190 11 635 1 549 555 1 328 186 Zhejiang 1 512 300 27 297 1 485 003 1 463 653 China total 61 899 940 18 658 742 43 241 198 52 -

Annual Report 2020 DEFINITIONS (CONTINUED)
CONTENTS I Definitions 2 II Corporate Information 8 III Company Profile 10 IV Chairman’s Statement 12 V Management’s Discussion and Analysis 19 VI Corporate Governance Report 45 VII Report of the Directors 75 VIII Profile of Directors, Supervisors, Senior Management and Employees 99 IX Report of the Supervisory Committee 112 X Independent Auditor's Report 117 XI Consolidated Statement of Profit or Loss and Other Comprehensive Income 123 XII Consolidated Statement of Financial Position 125 XIII Consolidated Statement of Changes in Equity 127 XIV Consolidated Statement of Cash Flows 129 XV Notes to Financial Statements 131 DEFINITIONS In this section, the definitions are presented in alphabetical order (A–Z). I. NAME OF EXPRESSWAY PROJECTS Airport Expressway Chengdu Airport Expressway Chengbei Exit Expressway Chengdu Chengbei Exit Expressway Chengle Expressway Sichuan Chengle (Chengdu-Leshan) Expressway Chengren Expressway Chengdu-Meishan (Renshou) Section of ChengZiLuChi (Chengdu-Zigong-Luzhou-Chishui) Expressway Chengya Expressway Sichuan Chengya (Chengdu-Ya’an) Expressway Chengyu Expressway Chengyu (Chengdu-Chongqing) Expressway (Sichuan Section) Suiguang Expressway Sichuan Suiguang (Suining-Guang’an) Expressway Suixi Expressway Sichuan Suixi (Suining-Xichong) Expressway Tianqiong Expressway Chengdu Tianfu New Area to Qionglai Expressway 2 Sichuan Expressway Company Limited Annual Report 2020 DEFINITIONS (CONTINUED) II. BRANCHES, SUBSIDIARIES AND PRINCIPAL INVESTED COMPANIES Airport Expressway Company Chengdu Airport Expressway Company -

Yan Yang Curriculum Vitae
YAN YANG CURRICULUM VITAE Address Address: 1601 Maple St., Ed Annex Room 149, Carrollton, GA 30118 Phone: (678) 839-6133 Email: [email protected] Academic Achievement Degree School Years B.A. Sichuan University, China 1995 - 1999 M.A. Southwest Jiaotong University 2001 - 2004 Ph.D. Oklahoma State University 2005 - 2009 M.A. Thesis: Textual Analysis of English for Science and Technology Ph.D Dissertation: The Relationship between Attributional Styles of Cultural Awareness and Cultural Competence for Preservice Teachers Honors: 1. Preparing Future Faculty in Psychology Fellowship, Oklahoma State University 2. Research Travel Award, Oklahoma State University 3. Research Assistantship, Oklahoma State University 4. Teaching Assistantship, Oklahoma State University 5. Graduation with Distinction Award, Sichuan University 6. Education Scholarship of the United Laboratories, Sichuan University 7. Advanced Student Award of Athletics, Sichuan University 8. Prominent Three-Good Student Award, Sichuan University 9. Outstanding Leadership of the Students’ Union, Sichuan University Employment History 8/1/2020-Present Professor Department of Educational Technology and Foundations, University of West Georgia, Carrollton GA 8/1/2015-7/31/2020 Associate Professor Department of Educational Technology and Foundations, University of West Georgia, Carrollton GA 8/1/2011-7/31/2015 Assistant Professor Department of Educational Technology and Foundations, University of West Georgia, Carrollton GA 9/1/2009-7/30/2011 Assistant Professor Department of Educational Psychology and Leadership Studies, University of Texas at Brownsville, Brownsville TX 8/15/2005-5/30/2009 Instructor & Research Assistant School of Applied Health and Educational Psychology, Oklahoma State University, Stillwater OK 6/1/1999-6/30/2005 Assistant Professor School of Teaching English to Speakers of Other Languages (TESOL), Southwest Jiaotong University, Chengdu, Sichuan, China Refereed Publications 1. -

Historical Background of Wang Yang-Ming's Philosophy of Mind
Ping Dong Historical Background of Wang Yang-ming’s Philosophy of Mind From the Perspective of his Life Story Historical Background of Wang Yang-ming’s Philosophy of Mind Ping Dong Historical Background of Wang Yang-ming’s Philosophy of Mind From the Perspective of his Life Story Ping Dong Zhejiang University Hangzhou, Zhejiang, China Translated by Xiaolu Wang Liang Cai School of International Studies School of Foreign Language Studies Zhejiang University Ningbo Institute of Technology Hangzhou, Zhejiang, China Zhejiang University Ningbo, Zhejiang, China ISBN 978-981-15-3035-7 ISBN 978-981-15-3036-4 (eBook) https://doi.org/10.1007/978-981-15-3036-4 © The Editor(s) (if applicable) and The Author(s) 2020. This book is an open access publication. Open Access This book is licensed under the terms of the Creative Commons Attribution- NonCommercial-NoDerivatives 4.0 International License (http://creativecommons.org/licenses/by-nc- nd/4.0/), which permits any noncommercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if you modified the licensed material. You do not have permission under this license to share adapted material derived from this book or parts of it. The images or other third party material in this book are included in the book’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the book’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. -

Research on Development Strategy Oftourism Industry in the Southern
Journal of Service Science and Management, 2018, 11, 83-93 http://www.scirp.org/journal/jssm ISSN Online: 1940-9907 ISSN Print: 1940-9893 Research on Development Strategy of Tourism Industry in the Southern Sichuan Economic Zone Hongying Li School of Economics, Sichuan University of Science & Engineering, Zigong, China How to cite this paper: Li, H.Y. (2018) Abstract Research on Development Strategy of Tour- ism Industry in the Southern Sichuan Eco- The Southern Sichuan Economic Zone is one of the earliest explored districts nomic Zone. Journal of Service Science and in Sichuan Province and has gradually become a new polar of economic Management, 11, 83-93. growth. There are abundant tourism resources that ensure a strong momen- https://doi.org/10.4236/jssm.2018.111008 tum of economic development for the tourism industry; however, several Received: January 1, 2018 problems still exist and await urgent solution. The proposed strategy of tour- Accepted: February 10, 2018 ism industry development will contribute to moving the obstacles that hinders Published: February 13, 2018 the development of tourism industry and achieving the transformation, up- Copyright © 2018 by author and grading and frog-leaping development of tourism industry in the Southern Scientific Research Publishing Inc. Sichuan Economic Zone. This work is licensed under the Creative Commons Attribution International Keywords License (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/ Development Strategy, Tourism Industry, Regional Cooperation, Southern Open Access Sichuan Economic Zone 1. Introduction The Southern Sichuan area is one of the earliest explored districts in Sichuan Province, where has been found human remain about five to ten thousand years ago [1]. -

“The Wheels That Transformed the City: the Historical Development of Public Transportation Systems in Shanghai, 1843-1937”
“The Wheels that Transformed the City: The Historical Development of Public Transportation Systems in Shanghai, 1843-1937” A Ph.D Dissertation Presented to The Academic Faculty by Fang Zhou In Partial Fulfillment Of the Requirements for the Degree Doctor of Philosophy in the School of History, Techology, and Society Georgia Institute of Technology December 2010 “The Wheels that Transformed the City: The Historical Development of Public Transportation Systems in Shanghai, 1843-1937” Approved by Prof. Hanchao Lu, Advisor Prof. John Krige School of History, Technology, and Society School of History, Technology, and Society Georgia Institute of Technology Georgia Institute of Technology Prof. Steven Usselman Prof. Ronald Bayor School of History, Technology, and Society School of History, Technology, and Society Georgia Institute of Technology Georgia Institute of Technology Prof. Parks Coble Date Approved: May 11, 2010 Department of History University of Nebraska ACKNOWLEDGEMENTS There are many individuals whom I would like to thank during the research and completion of my dissertation. First, my greatest thanks and appreciation goes to my advisor, Prof. Hanchao Lu. Prof. Lu had guided and encouraged me from the beginning of my journey at Georgia Tech. Besides being one of the leading scholars of Chinese history, Prof. Lu has been the best mentor I could have had. I am forever indebted to Prof. Lu for his kindness, generosity, and support during the past five years. Prof. Lu‟s continued confidence in me even as I made numerous mistakes and shortcomings in my academic development had been extremely inspiring. Words cannot express how much gratitude I feel for having the opportunity to be mentored and advised by Prof.