Reproductive Ecology and Pollen Representation Among Neotropical
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Evolutionary Ecology of Pollination and Reproduction of Tropical Plants
TROPICAL BIOLOGY AND CONSERVATION MANAGEMENT - Vol. V - Evolutionary Ecology af Pollination and Reproduction of Tropical Plants - M. Quesada, F. Rosas, Y. Herrerias-Diego, R. Aguliar, J.A. Lobo and G. Sanchez-Montoya EVOLUTIONARY ECOLOGY OF POLLINATION AND REPRODUCTION OF TROPICAL PLANTS M. Quesada and F. Rosas Centro de Investigaciones en Ecosistemas, Universidad Nacional Autónoma de México, México. Y. Herrerias-Diego Universidad Michoacana de San Nicolás de Hidalgo, Michoacán, México. R. Aguilar IMBIV - UNC - CONICET, C.C. 495,(5000) Córdoba, Argentina J.A. Lobo Escuela de Biología, Universidad de Costa Rica G. Sanchez-Montoya Centro de Investigaciones en Ecosistemas, Universidad Nacional Autónoma de México, México. Keywords: Pollination, tropical plants, diversity, mating systems, gender, conservation. Contents 1. Introduction 1.1. The Life Cycle of Angiosperms 1.2. Overview of Angiosperm Diversity 2. Degree of specificity of pollination system 3. Diversity of pollination systems 3.1. Beetle Pollination (Cantharophily) 3.2. Lepidoptera 3.2.1. Butterfly Pollination (Psychophily) 3.2.2. Moth Pollination (Phalaenophily) 3.3. Hymenoptera 3.3.1. Bee PollinationUNESCO (Melittophily) – EOLSS 3.3.2. Wasps 3.4. Fly Pollination (Myophily and Sapromyophily) 3.5. Bird Pollination (Ornitophily) 3.6. Bat PollinationSAMPLE (Chiropterophily) CHAPTERS 3.7. Pollination by No-Flying Mammals 3.8. Wind Pollination (Anemophily) 3.9. Water Pollination (Hydrophily) 4. Reproductive systems of angiosperms 4.1. Strategies that Reduce Selfing and/or Promote Cross-Pollination. 4.2. Self Incompatibility Systems 4.2.1. Incidence of Self Incompatibility in Tropical Forest 4.3. The Evolution of Separated Sexes from Hermaphroditism 4.3.1. From Distyly to Dioecy ©Encyclopedia Of. Life Support Systems (EOLSS) TROPICAL BIOLOGY AND CONSERVATION MANAGEMENT - Vol. -

Pollination and Evolution of Plant and Insect Interaction JPP 2017; 6(3): 304-311 Received: 03-03-2017 Accepted: 04-04-2017 Showket a Dar, Gh
Journal of Pharmacognosy and Phytochemistry 2017; 6(3): 304-311 E-ISSN: 2278-4136 P-ISSN: 2349-8234 Pollination and evolution of plant and insect interaction JPP 2017; 6(3): 304-311 Received: 03-03-2017 Accepted: 04-04-2017 Showket A Dar, Gh. I Hassan, Bilal A Padder, Ab R Wani and Sajad H Showket A Dar Parey Sher-e-Kashmir University of Agricultural Science and Technology, Shalimar, Jammu Abstract and Kashmir-India Flowers exploit insects to achieve pollination; at the same time insects exploit flowers for food. Insects and flowers are a partnership. Each insect group has evolved different sets of mouthparts to exploit the Gh. I Hassan food that flowers provide. From the insects' point of view collecting nectar or pollen is rather like fitting Sher-e-Kashmir University of a key into a lock; the mouthparts of each species can only exploit flowers of a certain size and shape. Agricultural Science and This is why, to support insect diversity in our gardens, we need to plant a diversity of suitable flowers. It Technology, Shalimar, Jammu is definitely not a case of 'one size fits all'. While some insects are generalists and can exploit a wide and Kashmir-India range of flowers, others are specialists and are quite particular in their needs. In flowering plants, pollen grains germinate to form pollen tubes that transport male gametes (sperm cells) to the egg cell in the Bilal A Padder embryo sac during sexual reproduction. Pollen tube biology is complex, presenting parallels with axon Sher-e-Kashmir University of guidance and moving cell systems in animals. -

Pollination of Cultivated Plants in the Tropics 111 Rrun.-Co Lcfcnow!Cdgmencle
ISSN 1010-1365 0 AGRICULTURAL Pollination of SERVICES cultivated plants BUL IN in the tropics 118 Food and Agriculture Organization of the United Nations FAO 6-lina AGRICULTUTZ4U. ionof SERNES cultivated plans in tetropics Edited by David W. Roubik Smithsonian Tropical Research Institute Balboa, Panama Food and Agriculture Organization of the United Nations F'Ø Rome, 1995 The designations employed and the presentation of material in this publication do not imply the expression of any opinion whatsoever on the part of the Food and Agriculture Organization of the United Nations concerning the legal status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries. M-11 ISBN 92-5-103659-4 All rights reserved. No part of this publication may be reproduced, stored in a retrieval system, or transmitted in any form or by any means, electronic, mechanical, photocopying or otherwise, without the prior permission of the copyright owner. Applications for such permission, with a statement of the purpose and extent of the reproduction, should be addressed to the Director, Publications Division, Food and Agriculture Organization of the United Nations, Viale delle Terme di Caracalla, 00100 Rome, Italy. FAO 1995 PlELi. uion are ted PlauAr David W. Roubilli (edita Footli-anal ISgt-iieulture Organization of the Untled Nations Contributors Marco Accorti Makhdzir Mardan Istituto Sperimentale per la Zoologia Agraria Universiti Pertanian Malaysia Cascine del Ricci° Malaysian Bee Research Development Team 50125 Firenze, Italy 43400 Serdang, Selangor, Malaysia Stephen L. Buchmann John K. S. Mbaya United States Department of Agriculture National Beekeeping Station Carl Hayden Bee Research Center P. -

Lecture 4: ROLE of HONEY BEES in CROSS POLLINATION - THEIR EXPLOITATION - CASE STUDIES with SELECTED CROPS
Lecture 4: ROLE OF HONEY BEES IN CROSS POLLINATION - THEIR EXPLOITATION - CASE STUDIES WITH SELECTED CROPS For SEXUAL reproduction in flowering plants transfer of anther to stigma is essential - Pollination Self pollination Transfer to sligma of same plant No external agents are involved Cross pollination Transfer pollen from one plant to stigma of another plant External agents are involved External agents involved in pollination A. Abiotic agents a. Wind (Anemophily) Wind carries pollen from one plant to another Flowers are small, inconspicuous, unattractive Pollen are dry and light in weight Stigma feathery with large surface area eg: Maize, barley, wheat, sugarcane b. Water (Hydrophily) Water carries pollen from one plant to other B. Biotic agents Bird, bat and insects are important biotic agents Among insects honey bees play major role Honey bees and flowering plants have coevolved In insect pollinated plants, flowers are large, brightly colour, distinct fragrance, presence of nectar and sticky pollen True honeybees (Apis spp.) - Most valuable pollinators of commercial crop Qualities of honeybees which make them good pollinators 1. Body covered with hairs and have structural adaptation for carrying nectar and pollen. 2. Bees - Not injurious to plants 3. Adult and larva feed on nectar and pollen - Available in plenty 4. Superior pollinators - Since store pollen and nectar for future use 5. No diapause - Need pollen throughout year 6. Body size and probascis length - Suitable for many crops 7. Pollinate wide variety of crops 8. Forage -

Aerobiological Investigation and in Vitro Studies of Pollen Grains From
ORIGINAL ARTICLE Aerobiological Investigation and In Vitro Studies of Pollen Grains From 2 Dominant Avenue Trees in Kolkata, India J Mandal,1 I Roy,2 S Chatterjee,2 S Gupta-Bhattacharya1 1Division of Palynology and Environmental Biology, Department of Botany, Bose Institute, Kolkata, India 2Allergy Department, Institute of Child Health, Kolkata, India ■ Abstract Background: Peltophorum pterocarpum and Delonix regia are dominant avenue trees in the city of Kolkata in India. They are well adapted to the humid tropical climate and also grow commonly in different parts of the country. Their pollen grains are reported to be airborne. Objective: The aim of this study was to conduct an aerobiological survey in Kolkata to determine the concentration and seasonal periodicity of pollen grains from P pterocarpum and D regia and to analyze the meteorological factors responsible for their levels in the atmosphere. In addition, we analyzed the prevalence of sensitization due to these grains among patients with seasonal respiratory allergy. Methods: An aerobiological survey was conducted with a volumetric Burkard sampler from 2004 to 2006. Correlations between meteorological parameters and pollen grain concentrations were assessed by Spearman correlation test. The protein profi le of the pollen extracts was studied by sodium dodecyl sulfate polyacrylamide gel electrophoresis. Finally, the allergenic potential of the pollen extracts was evaluated in patients with respiratory allergy by skin prick test, immunoglobulin (Ig) E enzyme-linked immunosorbent assay, and IgE immunoblotting. Results: P pterocarpum and D regia pollen grains occur from March to June and April to July, respectively. The pollen concentrations showed statistically signifi cant positive correlations with maximum temperature and wind speed. -
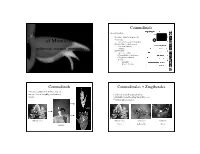
Diversity and Evolution of Monocots
Commelinids 4 main groups: Diversity and Evolution • Acorales - sister to all monocots • Alismatids of Monocots – inc. Aroids - jack in the pulpit • Lilioids (lilies, orchids, yams) – non-monophyletic . spiderworts, bananas, pineapples . – petaloid • Commelinids – Arecales – palms – Commelinales – spiderwort – Zingiberales –banana – Poales – pineapple – grasses & sedges Commelinids Commelinales + Zingiberales • theme: reduction of flower, loss of nectar, loss of zoophily, evolution of • 2 closely related tropical orders bracts • primarily nectar bearing but with losses • bracted inflorescences grass pickeral weed pickeral weed spiderwort heliconia nectar pollen only bracts rapatead bromeliad Commelinaceae - spiderwort Commelinaceae - spiderwort Family of small herbs with succulent stems, stems jointed; leaves sheathing. Family does not produce Inflorescence often bracted nectar, but showy flowers for insect pollen gathering. Rhoeo - Moses in a cradle Commelina erecta - Erect dayflower Tradescantia ohiensis - spiderwort Tradescantia ohiensis - spiderwort Commelinaceae - spiderwort Commelinaceae - spiderwort Flowers actinomorphic or • species rich in pantropics, CA 3 CO 3 A 6 G (3) zygomorphic especially Africa • floral diversity is enormous Commelina communis - day flower Tradescantia ohiensis - spiderwort Pontederiaceae - pickerel weed Pontederiaceae - pickerel weed Aquatic family of emergents or floaters. Pickerel weed has glossy heart-shaped leaves, Water hyacinth (Eichhornia) from superficially like Sagittaria but without net venation. -

Pollination Syndrome Table Pollinator Flower Characteristics Nectar Color Shape Odor Bloom Time Pollen Guides Bat
POLLINATION SYNDROME TABLE POLLINATOR FLOWER CHARACTERISTICS NECTAR COLOR SHAPE ODOR BLOOM TIME POLLEN GUIDES BAT Strong, musty, None Ample fruity Funnel or bowl shaped BEE Fresh, mild, Limited; often pleasant sticky, scented UV Closed lip with hori- zontal landing platform BUTTERFLY Faint but fresh Limited Composite with long, narrow tubes and landing platform HUMMINGBIRD None None Limited Tubular, hangs down- ward or sideways BATS Saguaro Agave Pollinator Characteristics: Color vision in white, green, purple range Good sense of smell, especially strong fragrances Large tongue to suck up hidden nectar Hairy head and body Good flyer Active during the night Flower Characteristics: BLOOM NECTAR COLOR SHAPE ODOR POLLEN TIME GUIDES Funnel or White or Strong, bowl shaped, Nighttime None Ample pale green high above musty, fruity ground Pollinated plants in Tohono Chul: Saguaro, Organ Pipe, Agave BEES Desert Willow Prickly Pear Mexican Sunflower Salvia Pollinator Characteristics: Large, compound eyes Good color vision in ultra-violet (UV), yellow, blue and green range Good sense of smell “Pack” pollen on hairy underside and between hairy legs to carry it back to nests to feed larvae Active during the day Flower Characteristics: BLOOM NECTAR COLOR SHAPE ODOR POLLEN TIME GUIDES Closed lip Bright white, with Limited; Fresh, mild, yellow, blue or horizontal Daytime Present often sticky, pleasant UV landing scented platform Pollinated plants in Tohono Chul: Carpenter bees: Desert Senna, Ocotillo (cheating), Squash, Tomatoes Bumblebees: -

Identification and Comparison of the Pollinators for the Purple-Fringed
University of Tennessee, Knoxville TRACE: Tennessee Research and Creative Exchange Supervised Undergraduate Student Research Chancellor’s Honors Program Projects and Creative Work Fall 1-2006 Identification and Comparison of the ollinatP ors for the Purple- fringed Orchids Platanthera psycodes and P.grandiflora John Richard Evans University of Tennessee-Knoxville Follow this and additional works at: https://trace.tennessee.edu/utk_chanhonoproj Recommended Citation Evans, John Richard, "Identification and Comparison of the ollinatP ors for the Purple-fringed Orchids Platanthera psycodes and P.grandiflora" (2006). Chancellor’s Honors Program Projects. https://trace.tennessee.edu/utk_chanhonoproj/953 This is brought to you for free and open access by the Supervised Undergraduate Student Research and Creative Work at TRACE: Tennessee Research and Creative Exchange. It has been accepted for inclusion in Chancellor’s Honors Program Projects by an authorized administrator of TRACE: Tennessee Research and Creative Exchange. For more information, please contact [email protected]. Identification and Comparison of the Pollinators for the Purple-fringed Orchids Platanthera psycodes and P. grandijlora John R. Evans Department of Ecology and Evolutionary Biology The University of Tennessee Advised by John A. Skinner Professor, Department of Entomology and Plant Pathology The University of Tennessee Abstract The pollination ecologies of the two purple-fringed orchids, Platanthera psycodes and P. grandiflora, are compared to test the prediction that, despite an extraordinary similarity in appearance, internal differences in the position and shape of fertile structures contributes to a distinct difference in their respective pollination ecologies. Field observations and experiments conducted during the 2003 flowering season in Great Smoky Mountains National Park revealed a number of effective pollen vectors not previously documented for P. -

Buzzing Bees and the Evolution of Sexual Floral Dimorphism in Australian Spiny Solanum
BUZZING BEES AND THE EVOLUTION OF SEXUAL FLORAL DIMORPHISM IN AUSTRALIAN SPINY SOLANUM ARTHUR SELWYN MARK School of Agriculture Food & Wine The University of Adelaide This thesis is submitted in fulfillment of the degree of Doctor of Philosophy June2014 1 2 Table of Contents List of Tables........................................................................................................... 6 List of Figures ......................................................................................................... 7 List of Boxes ......................................................................................................... 10 Abstract ................................................................................................................. 11 Declaration ............................................................................................................ 14 Acknowledgements ............................................................................................... 15 Chapter One - Introduction ................................................................................... 18 Floral structures for animal pollination .......................................................... 18 Specialisation in pollination .................................................................... 19 Specialisation in unisexual species ......................................................... 19 Australian Solanum species and their floral structures .................................. 21 Floral dimorphisms ................................................................................ -

Are Flower-Visiting Ants Mutualists Or Antagonists?Astudy in a Gynodioecious Wild Strawberry1
American Journal of Botany 92(5): 891±895. 2005. ARE FLOWER-VISITING ANTS MUTUALISTS OR ANTAGONISTS?ASTUDY IN A GYNODIOECIOUS WILD STRAWBERRY1 TIA-LYNN ASHMAN2 AND EMILEY A. KING Department of Biological Sciences, University of Pittsburgh, Pittsburgh, Pennsylvania 15260 USA; and Pymatuning Laboratory of Ecology, Linesville, Pennsylvania 16424 USA Ants are common ¯ower visitors, but their effects on plant reproductive ®tness have not often been assessed. Flower-visiting ants were studied to determine whether they are antagonists or mutualists and whether they could in¯uence ¯oral or breeding system evolution in gynodioecious wild strawberry (Fragaria virginiana). Ant and ¯ying pollinator (bees/¯ies) access to plants was manip- ulated, and visitation, fruit, and seed set were assessed. Ants visited ¯owers of hermaphrodites more often than those of females when bees and ¯ies were excluded, but visited the sex morphs equally when they were present. Insect class did not in¯uence fruit or seed set of hermaphrodites. In contrast, ants had both positive and negative effects on seed set in females. Females visited only by ants had 90% of the seed set of those visited only by bees/¯ies, and their seed set increased with ant visitation. The spatial pattern of seed set, however, suggests that ants may also damage pistils. Lastly, in contrast to bees and ¯ies, ants failed to increase visitation with ¯oral display size, suggesting that ant presence at ¯owers could reduce selection on this attractive trait. Findings suggest that when in high abundance, ¯ower-visiting ants could affect breeding system and ¯oral evolution in this gynodioecious plant. Key words: ant±plant interaction; ant pollination; ¯oral evolution; Fragaria virginiana; gynodioecy; pollination; Rosaceae; sexual dimorphism. -

Pollen Limitation and Flower Abortion in a Wind-Pollinated, Masting Tree
Notes Ecology, 96(2), 2015, pp. 587–593 Ó 2015 by the Ecological Society of America Pollen limitation and flower abortion in a wind-pollinated, masting tree 1,2,5 1,3 4 1 IAN S. PEARSE, WALTER D. KOENIG, KYLE A. FUNK, AND MARIO B. PESENDORFER 1Cornell Lab of Ornithology, 159 Sapsucker Woods Road, Ithaca, New York 14850 USA 2Illinois Natural History Survey, 1816 S. Oaks Street, Champaign, Illinois 61820 USA 3Department of Neurobiology and Behavior, Cornell University, Ithaca, New York 14853 USA 4School of Biological Sciences, University of Nebraska, Lincoln, Nebraska 68588 USA Abstract. Pollen limitation is a key assumption of theories that explain mast seeding, which is common among wind-pollinated and woody plants. In particular, the pollen coupling hypothesis and pollination Moran effect hypothesis assume pollen limitation as a factor that synchronizes seed crops across individuals. The existence of pollen limitation has not, however, been unambiguously demonstrated in wind-pollinated, masting trees. We conducted a two-year pollen supplementation experiment on a masting oak species, Quercus lobata. Supplemental pollen increased acorn set in one year but not in the other, supporting the importance of pollen coupling and pollination Moran effect models of mast seeding. We also tracked the fate of female flowers over five years and found that the vast majority of flowers were aborted for reasons unrelated to pollination, even in the presence of excess pollen. Pollen limitation can reduce annual seed set in a wind-pollinated tree, but factors other than pollen limitation cause the majority of flower abortion. Key words: anemophily; flower abortion; perennial plants; pollen; seed set; wind pollination. -

Pollination in Brazilian Syngonanthus (Eriocaulaceae) Species: Evidence for Entomophily Instead of Anemophily
Annals of Botany 96: 387–397, 2005 doi:10.1093/aob/mci191, available online at www.aob.oupjournals.org Pollination in Brazilian Syngonanthus (Eriocaulaceae) Species: Evidence for Entomophily Instead of Anemophily CARLIANNE O. C. RAMOS*, EDUARDO L. BORBA* and LI´GIA S. FUNCH Departamento de Cieˆncias Biolo´gicas, Laborato´rio de Taxonomia Vegetal, Universidade Estadual de Feira de Santana, Rodovia BR 116, km 03, Feira de Santana, BA, 44031-460, Brazil Received: 17 February 2005 Returned for revision: 1 April 2005 Accepted: 7 May 2005 Published electronically: 20 June 2005 Background and Aims The reproductive biology of Syngonanthus mucugensis and S. curralensis (Eriocaulaceae) was studied in areas of ‘campo rupestre’ vegetation in the Chapada Diamantina, north-eastern Brazil. These species are herbaceous and the individuals have a grouped distribution. Their leaves are united in a rosette, and their inflorescence is monoecious, of the capitulum type. The staminate and pistillate rings mature in a centripetal manner on the capitulum. Methods A field study was conducted, including observations concerning the morphology and biology of the flowers, fruit development, insect visits and anemophily, in both S. mucugensis and S. curralensis. Experimental pollinations were also carried out to study the mating systems of S. mucugensis. Key Results Both species flower from June to August. The staminate cycle lasts approx. 7 d, and the pistillate cycle from 3 to 4 d, with no temporal overlap between them on the same capitulum. The pollen viability of S. mucugensis was 88Á6 %, and 92Á5 % for S. curralensis. The inflorescences of both species demonstrated ultraviolet absorbance, and a sweet odour was detected during both the staminate and pistillate phases.