Territorial Signalling: a New Hypothesis to Explain Frequent Copulation in Raptorial Birds
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Birds (Aves) of Oromia, Ethiopia – an Annotated Checklist
European Journal of Taxonomy 306: 1–69 ISSN 2118-9773 https://doi.org/10.5852/ejt.2017.306 www.europeanjournaloftaxonomy.eu 2017 · Gedeon K. et al. This work is licensed under a Creative Commons Attribution 3.0 License. Monograph urn:lsid:zoobank.org:pub:A32EAE51-9051-458A-81DD-8EA921901CDC The birds (Aves) of Oromia, Ethiopia – an annotated checklist Kai GEDEON 1,*, Chemere ZEWDIE 2 & Till TÖPFER 3 1 Saxon Ornithologists’ Society, P.O. Box 1129, 09331 Hohenstein-Ernstthal, Germany. 2 Oromia Forest and Wildlife Enterprise, P.O. Box 1075, Debre Zeit, Ethiopia. 3 Zoological Research Museum Alexander Koenig, Centre for Taxonomy and Evolutionary Research, Adenauerallee 160, 53113 Bonn, Germany. * Corresponding author: [email protected] 2 Email: [email protected] 3 Email: [email protected] 1 urn:lsid:zoobank.org:author:F46B3F50-41E2-4629-9951-778F69A5BBA2 2 urn:lsid:zoobank.org:author:F59FEDB3-627A-4D52-A6CB-4F26846C0FC5 3 urn:lsid:zoobank.org:author:A87BE9B4-8FC6-4E11-8DB4-BDBB3CFBBEAA Abstract. Oromia is the largest National Regional State of Ethiopia. Here we present the first comprehensive checklist of its birds. A total of 804 bird species has been recorded, 601 of them confirmed (443) or assumed (158) to be breeding birds. At least 561 are all-year residents (and 31 more potentially so), at least 73 are Afrotropical migrants and visitors (and 44 more potentially so), and 184 are Palaearctic migrants and visitors (and eight more potentially so). Three species are endemic to Oromia, 18 to Ethiopia and 43 to the Horn of Africa. 170 Oromia bird species are biome restricted: 57 to the Afrotropical Highlands biome, 95 to the Somali-Masai biome, and 18 to the Sudan-Guinea Savanna biome. -

South Africa: Magoebaskloof and Kruger National Park Custom Tour Trip Report
SOUTH AFRICA: MAGOEBASKLOOF AND KRUGER NATIONAL PARK CUSTOM TOUR TRIP REPORT 24 February – 2 March 2019 By Jason Boyce This Verreaux’s Eagle-Owl showed nicely one late afternoon, puffing up his throat and neck when calling www.birdingecotours.com [email protected] 2 | TRIP REPORT South Africa: Magoebaskloof and Kruger National Park February 2019 Overview It’s common knowledge that South Africa has very much to offer as a birding destination, and the memory of this trip echoes those sentiments. With an itinerary set in one of South Africa’s premier birding provinces, the Limpopo Province, we were getting ready for a birding extravaganza. The forests of Magoebaskloof would be our first stop, spending a day and a half in the area and targeting forest special after forest special as well as tricky range-restricted species such as Short-clawed Lark and Gurney’s Sugarbird. Afterwards we would descend the eastern escarpment and head into Kruger National Park, where we would make our way to the northern sections. These included Punda Maria, Pafuri, and the Makuleke Concession – a mouthwatering birding itinerary that was sure to deliver. A pair of Woodland Kingfishers in the fever tree forest along the Limpopo River Detailed Report Day 1, 24th February 2019 – Transfer to Magoebaskloof We set out from Johannesburg after breakfast on a clear Sunday morning. The drive to Polokwane took us just over three hours. A number of birds along the way started our trip list; these included Hadada Ibis, Yellow-billed Kite, Southern Black Flycatcher, Village Weaver, and a few brilliant European Bee-eaters. -

A New Female-Like Morph of Juvenile Male Levant Sparrowhawk (Accipiter Brevipes) – Sexual Mimicry to Avoid Intra-Specific Predation?
EUROPEAN JOURNALEUROPEAN OF ECOLOGY JOURNAL OF ECOLOGY EJE 2015, 1(1): 64-67, doi: 10.1515/eje-2015-0008 A new female-like morph of juvenile male Levant Sparrowhawk (Accipiter brevipes) – sexual mimicry to avoid intra-specific predation? Reuven Yosef1, Lorenzo Fornasari2 1 Ben Gurion University ABSTRACT - Eilat Campus, P. O. Box In migrant Levant Sparrowhawk (Accipiter brevipes) at Eilat, Israel, we noted that juvenile males had two differ- 272, Eilat 88000, Israel ent morphs – the one described to date in literature; and a second, previously undescribed morph, with female- Corresponding Author: [email protected] like barring on the chest and flanks interspersed with tear-shaped elongated spots, giving an overall female-like appearance. Here we forward the hypothesis that explain the evolutionary consequences for the female-like 2 FaunaViva - Viale Sar- plumage of juvenile males as that of intra-specific sex mimicry developed to avoid intra-specific predation by ca, 78 - 20125 Milano, the larger females. Italy, e-mail: lorenzo. [email protected] KEYWORDS Levant sparrowhawk – intraspecific predation – avoidance – morph © 2015 Reuven Yosef, Lorenzo Fornasari This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivs license INTRODUCTION The Levant Sparrowhawk has dichromatism and re- Chromatic mimicry as a strategy to avoid inter-specific preda- versed sexual size dimorphism wherein the female is larger by tion, or to have a reproductive advantage, is well documented 9–10% than the male (Cramp & Simmons 1980; Clark & Yosef in many insect, amphibian and reptilian taxa (e.g.Gross & Char- 1997). The sexes also differ in colour, and the male has blue- nov 1980; Krebs & Davies 1987). -

Birds Versus Bats: Attack Strategies of Bat-Hunting Hawks, and the Dilution Effect of Swarming
Supplementary Information Accompanying: Birds versus bats: attack strategies of bat-hunting hawks, and the dilution effect of swarming Caroline H. Brighton1*, Lillias Zusi2, Kathryn McGowan2, Morgan Kinniry2, Laura N. Kloepper2*, Graham K. Taylor1 1Department of Zoology, University of Oxford, South Parks Road, Oxford, OX1 3PS, UK. 2Department of Biology, Saint Mary’s College, Notre Dame, IN 46556, USA. *Correspondence to: [email protected] This file contains: Figures S1-S2 Tables S1-S3 Supplementary References supporting Table S1 Legend for Data S1 and Code S1 Legend for Movie S1 Data S1 and Code S1 implementing the statistical analysis have been uploaded as Supporting Information. Movie S1 has been uploaded to figshare: https://doi.org/10.6084/m9.figshare.11823393 Figure S1. Video frames showing examples of attacks on lone bats and the column. (A,B) Attacks on the column of bats, defined as an attack on one or more bats within a cohesive group of individuals all flying in the same general direction. (C-E) Attacks on a lone bat (circled red), defined as an attack on an individual that appeared to be flying at least 1m from the edge of the column, and typically in a different direction to the swarm. (F) If an attack occurred in a volume containing many bats, but with no coherent flight direction, then this was also categorised as an attack on a lone bat, rather than as an attack on the swarm. Figure S2 Video frames used to estimate the proportion of bats meeting the criteria for classification as lone bats. -

Biodiversity Profile of Afghanistan
NEPA Biodiversity Profile of Afghanistan An Output of the National Capacity Needs Self-Assessment for Global Environment Management (NCSA) for Afghanistan June 2008 United Nations Environment Programme Post-Conflict and Disaster Management Branch First published in Kabul in 2008 by the United Nations Environment Programme. Copyright © 2008, United Nations Environment Programme. This publication may be reproduced in whole or in part and in any form for educational or non-profit purposes without special permission from the copyright holder, provided acknowledgement of the source is made. UNEP would appreciate receiving a copy of any publication that uses this publication as a source. No use of this publication may be made for resale or for any other commercial purpose whatsoever without prior permission in writing from the United Nations Environment Programme. United Nations Environment Programme Darulaman Kabul, Afghanistan Tel: +93 (0)799 382 571 E-mail: [email protected] Web: http://www.unep.org DISCLAIMER The contents of this volume do not necessarily reflect the views of UNEP, or contributory organizations. The designations employed and the presentations do not imply the expressions of any opinion whatsoever on the part of UNEP or contributory organizations concerning the legal status of any country, territory, city or area or its authority, or concerning the delimitation of its frontiers or boundaries. Unless otherwise credited, all the photos in this publication have been taken by the UNEP staff. Design and Layout: Rachel Dolores -

A Partial Post-Juvenile Molt and Transitional Plumage in the Shikra (Accipiter Badius) and Grey Frog Hawk ( a Ccipiter Soloensis)
THE JOURNAL OF RAPTOR RESEARCH A QUARTERLY PUBLICATION OF • THE RAPTOR RESEARCH FOUNDATION, INC. VOL. 34 DECEMBER 2000 NO. 4 J. RaptorRes. 34(4) :249-261 ¸ 2000 The Raptor Research Foundation, Inc. A PARTIAL POST-JUVENILE MOLT AND TRANSITIONAL PLUMAGE IN THE SHIKRA (ACCIPITER BADIUS) AND GREY FROG HAWK ( A CCIPITER SOLOENSIS) MARC HERREMANS AND MICHEL LOUETTE RoyalMuseum for CentralAfrica, Department Zoology, Leuvensesteenweg 13, B-3080 Tervuren,Belgium ABSTRACT.--Molthas been poorly studied in the Accipitridae. Examination of museum specimens showedthat there are three age-relatedplumages in the Shikra (Accipiterbadius) and Grey Frog Hawk (A. soloensis)similar to the pattern known in the Levant Sparrowhawk(A. brevipes).The juvenile plumage with its distinctively-spottedunderside is replacedby a transitionalpost-juvenile plumage during a partial contour molt between 4-10 mo of age. More feathers on the ventral side than on the dorsal side are replaced during this first contour molt, which is arrested at variousstages of incomplete feather replace- ment. Usually, a significantpart of the ventral pattern changesfrom spotted to barred, whereby the barring is on averagemore prominent than in adults. The early development of a transitional post- juvenile plumage might be related to early sex signaling.The adult plumage replacesthe transitional post-juvenileplumage during a completemolt at about one year of age. In the subspeciesA. b.poliopsis of the Shikra, which has almost no sexual dimorphism in the adult plumage, the transitional plumage is uncommon and very poorly developed. KEYWORDS: Shikra;Accipiter badius; Greyb?og Hawk; Accipiter soloensis;Levant Sparrowhawk; Accipiter brevipes;contour molt;, transitional post-juvenile plumage. Muda parcial postjuvenil y de transicionde plumaje en Accipiterbadius y Accipitersoloensis RES0MEN.--Lamuda ha sido poco esmdiada en las Accipitridae. -

Thailand Custom Tour 29 January -13 February, 2017
Tropical Birding Trip Report THAILAND JANUARY-FEBRUARY, 2017 Thailand custom tour 29 January -13 February, 2017 TOUR LEADER: Charley Hesse Report by Charley Hesse. Photos by Charley Hesse & Laurie Ross. All photos were taken on this tour When it comes to vacation destinations, Thailand has it all: great lodgings, delicious food, scenery, good roads, safety, value for money and friendly people. In addition to both its quantity & quality of birds, it is also one of the most rapidly evolving destinations for bird photography. There are of course perennial favourite locations that always produce quality birds, but year on year, Thailand comes up with more and more fantastic sites for bird photography. On this custom tour, we followed the tried and tested set departure itinerary and found an impressive 420 species of birds and 16 species of mammals. Some of the highlights included: Spoon-billed Sandpiper and Nordmann’s Greenshank around Pak Thale; Wreathed Hornbill, Long-tailed & Banded Broadbills inside Kaeng Krachan National Park; Rosy, Daurian & Spot-winged Starlings at a roost site just outside; Kalij Pheasant, Scaly-breasted & Bar-backed Partridges at a private photography blind nearby; Siamese Fireback and Great Hornbill plus Asian Elephant & Malayan Porcupine at Khao Yai National Park; countless water birds at Bueng Boraphet; a myriad of montane birds at Doi Inthanon; Giant Nuthatch at Doi Chiang Dao; Scarlet-faced Liocichla at Doi Ang Khang; Hume’s Pheasant & Spot-breasted Parrotbill at Doi Lang; Yellow-breasted Buntings at Baan Thaton; and Baikal Bush-Warbler & Ferruginous Duck at Chiang Saen. It was a truly unforgettable trip. www.tropicalbirding.com +1-409-515-9110 [email protected] Tropical Birding Trip Report THAILAND JANUARY-FEBRUARY, 2017 29th January – Bangkok to Laem Pak Bia After a morning arrival in Bangkok, we left the sprawling metropolis on the overhead highways, and soon had our first birding stop at the Khok Kham area of Samut Sakhon, the neighbouring city to Bangkok. -

Status and Distribution of Faunal Diversity in Kafa Afromontane Coffee Forest
Status and Distribution of Faunal Diversity in Kafa Afromontane Coffee Forest Leykun Abunie Berhan Submitted to PPP Project July 2008 Addis Ababa Contents Executive Summary .....................................................................................................................4 Introduction..................................................................................................................................6 Literature Review Related to Faunal Diversity and Management...............................................8 Macro Policies and Priorities......................................................................................................8 Environmental Protection Policy.................................................................................................8 Wildlife Development / Management Policy................................................................................9 Analysis of Wildlife Sector in Ethiopia ......................................................................................10 Physical and Ecological Description of the Study Area ............................................................14 Objective of the Present Study...................................................................................................16 Methodology ..............................................................................................................................17 General Approach......................................................................................................................17 -
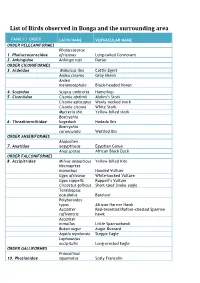
Birds Species in Bonga List
List of Birds observed in Bonga and the surrounding area FAMILY / ORDER LATIN NAME VERNACULAR NAME ORDER PELECANIFORMES Phalacrocorax 1. Phalacrocoracidae africanus Long-tailed Cormorant 2. Anhingidae Anhinga rufa Darter ORDER CICONIIFORMES 3. Ardeidae Bubulcus ibis Cattle Egret Ardea cinerea Grey Heron Ardea melanocephala Black-headed Heron 4. Scopidae Scopus umbretta Hamerkop 5. Ciconiidae Ciconia abdimii Abdim’s Stork Ciconia episcopus Wooly -necked stork Ciconia ciconia White Stork Mycteria ibis Yellow -billed stork Bostrychia 6. Threskiornithidae hagedash Hadada Ibis Bostrychia carunculata Wattled Ibis ORDER ANSERIFORMES Alopochen 7. Anatidae aegyptiacus Egyptian Goose Anas sparsa African Black Duck ORDER FALCONIFORMES 8. Accipitridae Milvus aegypticus Yellow -billed Kite Necrosyrtes monachus Hooded Vulture Gyps africanus White -backed Vulture Gyps ruppellii Ruppell’s Vulture Circaetus gallicus Short -toed Snake -eagle Terathopius ecaudatus Bateleur Polyboroides typus African Harrier Hawk Accipiter Red -breasted/Rufous -chested Sparrow rufiventris hawk Accipiter minullus Little Sparrowhawk Buteo augur Augur Buzzard Aquila nipalensis Steppe Eagle Lophoaetus occipitalis Long-crested Eagle ORDER GALLIFORMES Francolinus 10. Phasianidae squamatus Scaly Francolin FAMILY / ORDER LATIN NAME VERNACULAR NAME Francolinus castaneicollis Chestnut-napped Francolin ORDER GRUIFORMES Balearica 11. Gruidae pavonina Black Crowned Crane Bugeranus carunculatus Wattled crane Ruogetius 12. Rallidae rougetii Rouget’s Rail Podica 13. Heliornithidae senegalensis African Finfoot ORDER CHARADRIIFORMES 14. Scolopacidae Tringa ochropus Green Sandpiper Tringa hypolucos Common Sandpiper ORDER COLUMBIFORMES 15. Columbidae Columba guinea Speckled Pigeon Columba arquatrix African Olive Pigeon (Rameron Pigeon) Streptopelia lugens Dusky (Pink-breasted) Turtle Dove Streptopelia semitorquata Red-eyed Dove Turtur chalcospilos Emerald-spotted Wood Dove Turtur tympanistria Tambourine Dove ORDER CUCULIFORMES 17. Musophagidae Tauraco leucotis White -cheeked Turaco Centropus 18. -

Pakistan, an Exploration of Part of the World Whose Birds Are Poorly Known and Rarely Seen
THE WESTERN HIMALAYAS 19 - 31 JULY 2005 TOUR REPORT LEADER: DAVE FARROW GROUP MEMBERS: CHRISTA GLAUSER, WERNER MULLER, AVRIL McLEOD, BOB McLEOD and DAVE WEDD This was just the second Birdquest to the Western Himalayas of Pakistan, an exploration of part of the world whose birds are poorly known and rarely seen. Just a seven hour flight from London and only a four hour difference in time-zones, it seems like a short hop from Europe compared to many other birding destinations. We were able to enjoy some great birding in world-class scenery, recording 190 species, a birdlist rich in quality with many near-endemics and regional specialities such as Koklass Pheasant, Brown-fronted and Himalayan Woodpeckers, Pied, Lesser and Grey-bellied Cuckoos, Slaty-headed Parakeets, White-eyed and Himalayan Buzzards, Indian Golden Oriole, Tibetan Blackbird, Chestnut Thrush, Rusty-tailed Flycatcher, White-tailed Rubythroat, Himalayan Red-flanked and Golden Bush Robins, White-bellied Redstart, Kashmir and White-cheeked Nuthatches, Bar-tailed Treecreeper, Spot-winged, Rufous-naped, White-throated and the near-endemic White-cheeked Tits, Streak-throated Swallow, Indian Reed Warbler, Tickell’s Leaf, Sulphur-bellied, Lemon-rumped, Hume’s, Large-billed Leaf, Tytler’s Leaf and Western Crowned Warblers, Variegated Laughingthrush, Black-chinned Babbler, Upland and Rosy Pipits, Fire-fronted Serin, Grey-headed Goldfinch, Spectacled Finch, Pink-browed and Himalayan White-browed Rosefinches, Black-and-Yellow Grosbeak and White-capped and Chestnut-eared Buntings. Mammals were quite visible and we saw Common Goral, Kashmir Grey Langur, Golden Jackal, Long-tailed Marmot, Stoat and Kashmir Flying Squirrel. All this was easily accomplished in a nation that is remarkably friendly and we were well looked after, allowing us to have a safe, productive and comfortable visit. -

Resident Forest Bird Populations and Co- Management Impacts
CREL Knowledge and Impact Series – Report 1 Resident Forest Bird Populations and Co- Management Impacts Enam Ul Haque, Sayam U. Chowdhury, Samiul Mohsanin, Paul M. Thompson Climate-Resilient Ecosystems and Livelihoods (CREL) in association with Bangladesh bird club AID-388-A-12-00007 Dhaka August 2018 Resident Forest Bird Populations and Co-Management Impacts Enam Ul Haque, Sayam U. Chowdhury, Samiul Mohsanin, Paul M. Thompson Dhaka August 2018 Cover photo: Green-billed Malkoha (Sayam U. Chowdhury), above photo: Survey team ( Sayam U. Chowdhury) Climate-Resilient Ecosystems and Livelihoods (CREL) project House 13/B Road 54, Gulshan Dhaka 1212, Bangladesh Winrock International 2101 Riverfront Drive Little Rock, AR 72202-1748, USA Bangladesh bird club House 11, Road 4, Banani DOHS Dhaka 1206, Bangladesh Citation: Haque, E.U., Chowdhury, S.U., Mohsanin, S., Thompson, P. (2018). Resident Forest Bird Populations and Co-Management Impacts. CREL Technical Report No. 1. Climate-Resilient Ecosystems and Livelihoods (CREL) Project, Dhaka, Bangladesh. This publication is made possible by the generous support of the American people through the United States Agency for International Development (USAID). The contents of this document do not necessarily reflect the views of the USAID or the United States Government. CREL TECHNICAL REPORT 2 i FOREST BIRD SURVEYS TABLE OF CONTENTS Page 1 BACKGROUND 1-1 1.1 Introduction 1-1 1.2 Conceptual Framework 1-2 1.3 Capacity Building of Local Surveyors 1-2 2 METHODS 2-1 2.1 Species and Sites Monitored 2-1 2.2 Transect -

Black Wildebeest
SSOOUUTTHHEERRNN AAFFRRIICCAANN RRAAPPTTOORR CCOONNSSEERRVVAATTIIOONN STRATEGIC PLANNING WORKSHOP REPORT 23 – 25 March 2004 Gariep Dam, Free State, South Africa Hosted by: THE RAPTOR CONSERVATION GROUP OF THE ENDANGERED WILDLIFE TRUST Sponsored by: SA EAGLE INSURANCE COMPANY ESKOM In collaboration with: THE CONSERVATION BREEDING SPECIALIST GROUP SOUTHERN AFRICA (CBSG – SSC/IUCN) 0 SSOOUUTTHHEERRNN AAFFRRIICCAANN RRAAPPTTOORR CCOONNSSEERRVVAATTIIOONN STRATEGIC PLANNING WORKSHOP REPORT The Raptor Conservation Group wishes to thank Eskom and SA Eagle Insurance company for the sponsorship of this publication and the workshop. Evans, S.W., Jenkins, A., Anderson, M., van Zyl, A., le Roux, J., Oertel, T., Grafton, S., Bernitz Z., Whittington-Jones, C. and Friedmann Y. (editors). 2004. Southern African Raptor Conservation Strategic Plan. Conservation Breeding Specialist Group (SSC / IUCN). Endangered Wildlife Trust. 1 © Conservation Breeding Specialist Group (CBSG-SSC/IUCN) and the Endangered Wildlife Trust. The copyright of the report serves to protect the Conservation Breeding Specialist Group workshop process from any unauthorised use. The CBSG, SSC and IUCN encourage the convening of workshops and other fora for the consideration and analysis of issues related to conservation, and believe that reports of these meetings are most useful when broadly disseminated. The opinions and recommendations expressed in this report reflect the issues discussed and ideas expressed by the participants during the Southern African Raptor Conservation Strategic