Birds Versus Bats: Attack Strategies of Bat-Hunting Hawks, and the Dilution Effect of Swarming
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Mississippi Kite (Ictinia Mississippiensis)
Mississippi Kite (Ictinia mississippiensis) NMPIF level: Species Conservation Concern, Level 2 (SC2) NMPIF assessment score: 15 NM stewardship responsibility: Low National PIF status: No special status New Mexico BCRs: 16, 18, 35 Primary breeding habitat(s): Urban (southeast plains) Other habitats used: Agricultural, Middle Elevation Riparian Summary of Concern Mississippi Kite is a migratory raptor that has successfully colonized urban habitats (parks, golf courses, residential neighborhoods) in the western portion of its breeding range over the last several decades. Little is known about species ecology outside of the breeding season and, despite stable or increasing populations at the periphery of its range, it remains vulnerable due to its small population size. Associated Species Cooper’s Hawk, Ring-necked Pheasant, Mourning Dove, American Robin Distribution Mississippi Kite is erratically distributed across portions of the east and southeast, the southern Great Plains, and the southwest, west to central Arizona and south to northwest Chihuahua. It is most abundant in areas of the Gulf Coast, and in the Texas and Oklahoma panhandles. The species is a long- distance migrant, wintering in Argentina, Paraguay, and perhaps other locations in South America. In New Mexico, Mississippi Kite is most common in cites and towns of the southeast plains. It is also present in the Middle Rio Grande valley north to Corrales, and the Pecos River Valley north to Fort Sumner and possibly Puerto de Luna (Parker 1999, Parmeter et al. 2002). Ecology and Habitat Requirements Mississippi Kite occupies different habitats in different parts of its range, including mature hardwood forests in the southeast, rural woodlands in mixed and shortgrass prairie in the Great Plains, and mixed riparian woodlands in the southwest. -

Progress in the Development of an Eurasian-African Bird Migration Atlas
CONVENTION ON UNEP/CMS/COP13/Inf.20 MIGRATORY 10 February 2020 SPECIES Original: English 13th MEETING OF THE CONFERENCE OF THE PARTIES Gandhinagar, India, 17 - 22 February 2020 Agenda Item 25 PROGRESS IN THE DEVELOPMENT OF AN EURASIAN-AFRICAN BIRD MIGRATION ATLAS (Submitted by the European Union of Bird Ringing (EURING) and the Institute of Avian Research) Summary: The African-Eurasian Bird Migration Atlas is being developed under the auspices of CMS in the framework of a Global Animal Migration Atlas, of which it constitutes a module. The African-Eurasian Bird Migration Atlas is being developed and compiled by the European Union of Bird Ringing (EURING) under a Project Cooperation Agreement (PCA) between the CMS Secretariat and the Institute of Avian Research, acting on behalf of EURING. The development of the African-Eurasian Bird Migration Atlas is funded with the contribution granted by the Government of Italy under the Migratory Species Champion Programme. This information document includes a progress report on the development of the various components of the project. The project is expected to be completed in 2021. UNEP/CMS/COP13/Inf.20 Eurasian-African Bird Migration Atlas progress report February 2020 Stephen Baillie1, Franz Bairlein2, Wolfgang Fiedler3, Fernando Spina4, Kasper Thorup5, Sam Franks1, Dorian Moss1, Justin Walker1, Daniel Higgins1, Roberto Ambrosini6, Niccolò Fattorini6, Juan Arizaga7, Maite Laso7, Frédéric Jiguet8, Boris Nikolov9, Henk van der Jeugd10, Andy Musgrove1, Mark Hammond1 and William Skellorn1. A report to the Convention on Migratory Species from the European Union for Bird Ringing (EURING) and the Institite of Avian Research, Wilhelmshaven, Germany 1. British Trust for Ornithology, Thetford, IP24 2PU, UK 2. -
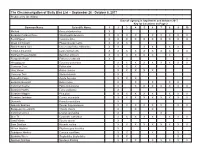
The Circumnavigation of Sicily Bird List -- September 26
The Circumnavigation of Sicily Bird List -- September 26 - October 8, 2017 Produced by Jim Wilson Date of sighting in September and October 2017 Key for Locations on Page 2 Common Name Scientific Name 1 2 3 4 5 6 7 8 9 10 Mallard Anas platyrhynchos X Eurasian Collared Dove Streptopelia decaocto X X X X X X X X X Feral Pigeon Columba livia X X X X X X X X X X Great Cormorant Phalacrocorax carbo X X X Black-headed Gull Chroicocephalus ridibundus X X X X X X Yellow-legged Gull Larus michahellis X X X X X X X X X X Northern House Martin Delichon urbicum X X X X X X X European Robin Erithacus rubecula X X Woodpigeon Columba palumbus X X X X X X X X X Common Coot Fulica atra X X X Grey Heron Ardea cinerea X X X X X Common Tern Sterna hirundo X Bonnelli's Eagle Aquila fasciata X X X Eurasian Buzzard Buteo buteo X X X X X Common Kestrel Falco tinnunculus X X X X X X X X Eurasian Hobby Falco subbuteo X Eurasian Magpie Pica pica X X X X X X X X Eurasian Jackdaw Corvus monedula X X X X X X X X X Dunnock Prunella modularis X Spanish Sparrow Passer hispaniolensis X X X X X X X X European Greenfinch Chloris chloris X X X Common Linnet Linaria cannabina X X Blue Tit Cyanistes caeruleus X X X X X X X X Sand Martin Riparia riparia X Barn Swallow Hirundo rustica X X X X X X X X X Willow Warbler Phylloscopus trochilus X Subalpine Warbler Curruca cantillans X X X X Eurasian Wren Troglodytes troglodytes X X X Spotless Starling Spotless Starling X X X X X X X X Common Name Scientific Name 1 2 3 4 5 6 7 8 9 10 Sardinian Warbler Curruca melanocephala X X X X -

HAVE the EGGS of the ORANGE&Hyphen;BREASTED
HAVE THE EGGS OF THE ORANGE-BREASTED FALCON (FALCO DEIROLEUCUS) BEEN DESCRIBED? by DouglasA. BoyceJr.' School of Natural Resources HumboldtState University Arcata, California 95521 and Lloyd F. Kiff WesternFoundation of VertebrateZoology 1100 Glendon Avenue LosAngeles, California 90024 The only publisheddescription of Orange-breastedFalcon (Falcodeiroleucus) eggs knownto us is that of Coltart (1952),who presenteddetails on two setsof eggssaid to be of thisspecies collected for G. D. Smookerin Trinidad.One of thesesets, a clutchof 3 takenon 28 March 1937in the Aripo Savannah,is now in the collectionof the West- ernFoundation of VertebrateZoology (WFVZ cat. no. 15,728);the other,a setof 2 col- lectedon 21 April 1930 in the Coroni Marshes,is in the collectionof the Zoological Museum,University of Helsinki,Finland (ZMUF cat. no. 15,721). The authenticityof theseeggs has been questioned by ffrench(1973) because of their smallsize compared to the bodysize of the species.The 3 eggsin the WFVZ collection measure40.6 x 34.7, 41.8 x 35.6, and 39.9 x 34.6 mm, and the 2 eggsin the ZMUF set measure 43.0 x 35.0 and 42.2 x 34.7 mm. All of these measurements fall within the rangegiven for eggsof the AplomadoFalcon (Falco femoralis) by Bent (1938) and Brownand Amadon(1968) and are only slightlylarger than the extrememeasurements knownfor eggsof the muchsmaller Bat Falcon(Falco rufigularis) (Brown and Amadon op cit., Kiff unpubl.data). Eggs of severalfalcon species, including one of Smooker's Trinidadeggs, are shownin Figurei to illustratetheir comparativesizes. Heinroth(1922) first demonstratedthe fundamentalrelationship between egg weight andbody weight in birds,and this was further refined by Huxley(1923-1924). -

A New Female-Like Morph of Juvenile Male Levant Sparrowhawk (Accipiter Brevipes) – Sexual Mimicry to Avoid Intra-Specific Predation?
EUROPEAN JOURNALEUROPEAN OF ECOLOGY JOURNAL OF ECOLOGY EJE 2015, 1(1): 64-67, doi: 10.1515/eje-2015-0008 A new female-like morph of juvenile male Levant Sparrowhawk (Accipiter brevipes) – sexual mimicry to avoid intra-specific predation? Reuven Yosef1, Lorenzo Fornasari2 1 Ben Gurion University ABSTRACT - Eilat Campus, P. O. Box In migrant Levant Sparrowhawk (Accipiter brevipes) at Eilat, Israel, we noted that juvenile males had two differ- 272, Eilat 88000, Israel ent morphs – the one described to date in literature; and a second, previously undescribed morph, with female- Corresponding Author: [email protected] like barring on the chest and flanks interspersed with tear-shaped elongated spots, giving an overall female-like appearance. Here we forward the hypothesis that explain the evolutionary consequences for the female-like 2 FaunaViva - Viale Sar- plumage of juvenile males as that of intra-specific sex mimicry developed to avoid intra-specific predation by ca, 78 - 20125 Milano, the larger females. Italy, e-mail: lorenzo. [email protected] KEYWORDS Levant sparrowhawk – intraspecific predation – avoidance – morph © 2015 Reuven Yosef, Lorenzo Fornasari This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivs license INTRODUCTION The Levant Sparrowhawk has dichromatism and re- Chromatic mimicry as a strategy to avoid inter-specific preda- versed sexual size dimorphism wherein the female is larger by tion, or to have a reproductive advantage, is well documented 9–10% than the male (Cramp & Simmons 1980; Clark & Yosef in many insect, amphibian and reptilian taxa (e.g.Gross & Char- 1997). The sexes also differ in colour, and the male has blue- nov 1980; Krebs & Davies 1987). -

Bird) Species List
Aves (Bird) Species List Higher Classification1 Kingdom: Animalia, Phyllum: Chordata, Class: Reptilia, Diapsida, Archosauria, Aves Order (O:) and Family (F:) English Name2 Scientific Name3 O: Tinamiformes (Tinamous) F: Tinamidae (Tinamous) Great Tinamou Tinamus major Highland Tinamou Nothocercus bonapartei O: Galliformes (Turkeys, Pheasants & Quail) F: Cracidae Black Guan Chamaepetes unicolor (Chachalacas, Guans & Curassows) Gray-headed Chachalaca Ortalis cinereiceps F: Odontophoridae (New World Quail) Black-breasted Wood-quail Odontophorus leucolaemus Buffy-crowned Wood-Partridge Dendrortyx leucophrys Marbled Wood-Quail Odontophorus gujanensis Spotted Wood-Quail Odontophorus guttatus O: Suliformes (Cormorants) F: Fregatidae (Frigatebirds) Magnificent Frigatebird Fregata magnificens O: Pelecaniformes (Pelicans, Tropicbirds & Allies) F: Ardeidae (Herons, Egrets & Bitterns) Cattle Egret Bubulcus ibis O: Charadriiformes (Sandpipers & Allies) F: Scolopacidae (Sandpipers) Spotted Sandpiper Actitis macularius O: Gruiformes (Cranes & Allies) F: Rallidae (Rails) Gray-Cowled Wood-Rail Aramides cajaneus O: Accipitriformes (Diurnal Birds of Prey) F: Cathartidae (Vultures & Condors) Black Vulture Coragyps atratus Turkey Vulture Cathartes aura F: Pandionidae (Osprey) Osprey Pandion haliaetus F: Accipitridae (Hawks, Eagles & Kites) Barred Hawk Morphnarchus princeps Broad-winged Hawk Buteo platypterus Double-toothed Kite Harpagus bidentatus Gray-headed Kite Leptodon cayanensis Northern Harrier Circus cyaneus Ornate Hawk-Eagle Spizaetus ornatus Red-tailed -

Syringeal Morphology and the Phylogeny of the Falconidae’
The Condor 96:127-140 Q The Cooper Ornithological Society 1994 SYRINGEAL MORPHOLOGY AND THE PHYLOGENY OF THE FALCONIDAE’ CAROLES.GRIFFITHS Departmentof Ornithology,American Museum of NaturalHistory and Departmentef Biology, City Collegeof City Universityof New York, Central Park West at 79th St., New York, NY 10024 Abstract. Variation in syringealmorphology was studied to resolve the relationshipsof representativesof all of the recognized genera of falcons, falconets, pygmy falcons, and caracarasin the family Falconidae. The phylogenyderived from thesedata establishesthree major cladeswithin the family: (1) the Polyborinae, containingDaptrius, Polyborus, Milvago and Phalcoboenus,the four genera of caracaras;(2) the Falconinae, consistingof the genus Falco, Polihierax (pygmy falcons),Spiziapteryx and Microhierax (falconets)and Herpetothe- res (Laughing Falcon); and (3) the genus Micrastur(forest falcons) comprising the third, basal clade. Two genera, Daptriusand Polihierax,are found to be polyphyletic. The phy- logeny inferred from these syringealdata do not support the current division of the family into two subfamilies. Key words: Falconidae;phylogeny; systematics; syrinx; falcons; caracaras. INTRODUCTION 1. The Polyborinae. This includes seven gen- Phylogenetic relationships form the basis for re- era: Daptrius, Milvago, Polyborus and Phalco- searchin comparative and evolutionary biology boenus(the caracaras),Micrastur (forest falcons), (Page1 and Harvey 1988, Gittleman and Luh Herpetotheres(Laughing Falcon) and Spiziapter- 1992). Patterns drawn from cladogramsprovide yx (Spot-winged Falconet). the blueprints for understanding biodiversity, 2. The Falconinae. This includes three genera: biogeography,behavior, and parasite-hostcospe- Falco, Polihierax (pygmy falcons) and Micro- ciation (Vane-Wright et al. 199 1, Mayden 1988, hierax (falconets). Page 1988, Coddington 1988) and are one of the Inclusion of the caracarasin the Polyborinae key ingredients for planning conservation strat- is not questioned (Sharpe 1874, Swann 1922, egies(Erwin 199 1, May 1990). -

Field Identification of Orange-Breasted and Bat Falcons
C O T IN G A 4 Orange-breasted and Bat falcons Field identification of Orange-breasted and Bat Falcons Steve N. G. Howell and Andrew Whittaker Introduction extent on chest of Bat Falcon); definite coarse The Bat Falcon Falco rufigularis is a wide buff barring on black vest (not fine white spread, small, generally quite common edging); and proportionately much stronger Neotropical falcon resident from Mexico to larger feet.” Other guides13,18 echo these fea South America, west of the Andes to Ecuador, tures, in particular the orange chest, buff east of the Andes to Brazil1. Over much of its barring on the narrower black chest band, and range the Bat Falcon is sympatric with the the proportionately larger feet of Orange larger and much rarer Orange-breasted Fal breasted. However, many Bat Falcons show con F. deiroleucus, which is resident from orangish to bright orange across the chest south-eastern Mexico to South America, east above the black vest (as cautioned by Ridgely of the Andes to Brazil1. These two species are & Gwynne16), the barring on the black vest can remarkably similar in plumage (Figure 1), but be whitish or buffy in both species, and judg the field identification of Orange-breasted, a ing the relative size of bill or feet on a lone near-threatened species6, is poorly treated in bird can be difficult. Becoming thoroughly many guides. Here we discuss characters that familiar with the structure and plumage vari can be used to help correctly identify Orange ation of the much commoner Bat Falcon is breasted Falcons. -

Biodiversity Profile of Afghanistan
NEPA Biodiversity Profile of Afghanistan An Output of the National Capacity Needs Self-Assessment for Global Environment Management (NCSA) for Afghanistan June 2008 United Nations Environment Programme Post-Conflict and Disaster Management Branch First published in Kabul in 2008 by the United Nations Environment Programme. Copyright © 2008, United Nations Environment Programme. This publication may be reproduced in whole or in part and in any form for educational or non-profit purposes without special permission from the copyright holder, provided acknowledgement of the source is made. UNEP would appreciate receiving a copy of any publication that uses this publication as a source. No use of this publication may be made for resale or for any other commercial purpose whatsoever without prior permission in writing from the United Nations Environment Programme. United Nations Environment Programme Darulaman Kabul, Afghanistan Tel: +93 (0)799 382 571 E-mail: [email protected] Web: http://www.unep.org DISCLAIMER The contents of this volume do not necessarily reflect the views of UNEP, or contributory organizations. The designations employed and the presentations do not imply the expressions of any opinion whatsoever on the part of UNEP or contributory organizations concerning the legal status of any country, territory, city or area or its authority, or concerning the delimitation of its frontiers or boundaries. Unless otherwise credited, all the photos in this publication have been taken by the UNEP staff. Design and Layout: Rachel Dolores -

The Birds of the Dar Es Salaam Area, Tanzania
Le Gerfaut, 77 : 205–258 (1987) BIRDS OF THE DAR ES SALAAM AREA, TANZANIA W.G. Harvey and KM. Howell INTRODUCTION Although the birds of other areas in Tanzania have been studied in detail, those of the coast near Dar es Salaam have received relatively little recent attention. Ruggles-Brise (1927) published a popular account of some species from Dar es Salaam, and Fuggles-Couchman (1939,1951, 1953, 1954, 1962) included the area in a series of papers of a wider scope. More recently there have been a few other stu dies dealing with particular localities (Gardiner and Gardiner 1971), habitats (Stuart and van der Willigen 1979; Howell 1981), or with individual species or groups (Harvey 1971–1975; Howell 1973, 1977). Britton (1978, 1981) has docu mented specimens collected in the area previous to 1967 by Anderson and others. The purpose of this paper is to draw together data from published reports, unpu blished records, museum specimens and our own observations on the frequency, habitat, distribution and breeding of the birds of the Dar es Salaam area, here defi ned as the portion of the mainland within a 64-km radius of Dar es Salaam, inclu ding the small islands just offshore (Fig. 1). It includes Dar es Salaam District and portions of two others, Kisarawe and Bagamoyo. Zanzibar has been omitted because its unusual avifauna has been reviewed (Pakenham 1979). Most of the mainland areas are readily accessible from Dar es Salaam by road and the small islands may be reached by boat. The geography of the area is described in Sutton (1970). -

A Partial Post-Juvenile Molt and Transitional Plumage in the Shikra (Accipiter Badius) and Grey Frog Hawk ( a Ccipiter Soloensis)
THE JOURNAL OF RAPTOR RESEARCH A QUARTERLY PUBLICATION OF • THE RAPTOR RESEARCH FOUNDATION, INC. VOL. 34 DECEMBER 2000 NO. 4 J. RaptorRes. 34(4) :249-261 ¸ 2000 The Raptor Research Foundation, Inc. A PARTIAL POST-JUVENILE MOLT AND TRANSITIONAL PLUMAGE IN THE SHIKRA (ACCIPITER BADIUS) AND GREY FROG HAWK ( A CCIPITER SOLOENSIS) MARC HERREMANS AND MICHEL LOUETTE RoyalMuseum for CentralAfrica, Department Zoology, Leuvensesteenweg 13, B-3080 Tervuren,Belgium ABSTRACT.--Molthas been poorly studied in the Accipitridae. Examination of museum specimens showedthat there are three age-relatedplumages in the Shikra (Accipiterbadius) and Grey Frog Hawk (A. soloensis)similar to the pattern known in the Levant Sparrowhawk(A. brevipes).The juvenile plumage with its distinctively-spottedunderside is replacedby a transitionalpost-juvenile plumage during a partial contour molt between 4-10 mo of age. More feathers on the ventral side than on the dorsal side are replaced during this first contour molt, which is arrested at variousstages of incomplete feather replace- ment. Usually, a significantpart of the ventral pattern changesfrom spotted to barred, whereby the barring is on averagemore prominent than in adults. The early development of a transitional post- juvenile plumage might be related to early sex signaling.The adult plumage replacesthe transitional post-juvenileplumage during a completemolt at about one year of age. In the subspeciesA. b.poliopsis of the Shikra, which has almost no sexual dimorphism in the adult plumage, the transitional plumage is uncommon and very poorly developed. KEYWORDS: Shikra;Accipiter badius; Greyb?og Hawk; Accipiter soloensis;Levant Sparrowhawk; Accipiter brevipes;contour molt;, transitional post-juvenile plumage. Muda parcial postjuvenil y de transicionde plumaje en Accipiterbadius y Accipitersoloensis RES0MEN.--Lamuda ha sido poco esmdiada en las Accipitridae. -

Introduction to Tropical Biodiversity, October 14-22, 2019
INTRODUCTION TO TROPICAL BIODIVERSITY October 14-22, 2019 Sponsored by the Canopy Family and Naturalist Journeys Participants: Linda, Maria, Andrew, Pete, Ellen, Hsin-Chih, KC and Cathie Guest Scientists: Drs. Carol Simon and Howard Topoff Canopy Guides: Igua Jimenez, Dr. Rosa Quesada, Danilo Rodriguez and Danilo Rodriguez, Jr. Prepared by Carol Simon and Howard Topoff Our group spent four nights in the Panamanian lowlands at the Canopy Tower and another four in cloud forest at the Canopy Lodge. In very different habitats, and at different elevations, conditions were optimal for us to see a great variety of birds, butterflies and other insects and arachnids, frogs, lizards and mammals. In general we were in the field twice a day, and added several night excursions. We also visited cultural centers such as the El Valle Market, an Embera Village, the Miraflores Locks on the Panama Canal and the BioMuseo in Panama City, which celebrates Panamanian biodiversity. The trip was enhanced by almost daily lectures by our guest scientists. Geoffroy’s Tamarin, Canopy Tower, Photo by Howard Topoff Hot Lips, Canopy Tower, Photo by Howard Topoff Itinerary: October 14: Arrival and Orientation at Canopy Tower October 15: Plantation Road, Summit Gardens and local night drive October 16: Pipeline Road and BioMuseo October 17: Gatun Lake boat ride, Emberra village, Summit Ponds and Old Gamboa Road October 18: Gamboa Resort grounds, Miraflores Locks, transfer from Canopy Tower to Canopy Lodge October 19: La Mesa and Las Minas Roads, Canopy Adventure, Para Iguana