Kissonerga Mosphilia
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Cyprus Authentic Route 2
Cyprus Authentic Route 2 Safety Driving in Cyprus Comfort Rural Accommodation Tips Useful Information Only DIGITAL Version A Village Life Larnaka • Livadia • Kellia • Troulloi • Avdellero • Athienou • Petrofani • Lympia • Ancient Idalion • Alampra • Mosfiloti • Kornos • Pyrga • Stavrovouni • Kofinou • Psematismenos • Maroni • Agios Theodoros • Alaminos • Mazotos • Kiti • Hala Sultan Tekke • Larnaka Route 2 Larnaka – Livadia – Kellia – Troulloi – Avdellero – Athienou – Petrofani – Lympia - Ancient Idalion – Alampra – Mosfiloti – Kornos – Pyrga – Stavrovouni – Kofinou – Psematismenos – Maroni – Agios Theodoros – Alaminos – Mazotos – Kiti – Hala Sultan Tekke – Larnaka Margo Agios Arsos Pyrogi Spyridon Agios Tremetousia Tseri Golgoi Sozomenos Melouseia Athienou Potamia Pergamos Petrofani Troulloi Margi Nisou Dali Pera Louroukina Avdellero Pyla Chorio Idalion Kotsiatis Lympia Alampra Agia Voroklini Varvara Agios Kellia Antonios Kochi Mathiatis Sia Aradippou Mosfiloti Agia Livadia Psevdas Anna Ε4 Kalo Chorio Port Kition Kornos Chapelle Delikipos Pyrga Royal LARNAKA Marina Salt LARNAKA BAY Lake Hala Sultan Stavrovouni Klavdia Tekkesi Dromolaxia- Dipotamos Meneou Larnaka Dam Kiti Dam International Alethriko Airport Tersefanou Anglisides Panagia Kivisili Menogeia Kiti Aggeloktisti Perivolia Aplanta Softades Skarinou Kofinou Anafotida Choirokoitia Alaminos Mazotos Cape Kiti Choirokoitia Agios Theodoros Tochni Psematismenos Maroni scale 1:300,000 0 1 2 4 6 8 10 Kilometers Zygi AMMOCHOSTOS Prepared by Lands and Surveys Department, Ministry of Interior, -

Mfi Id Name Address Postal City Head Office
MFI ID NAME ADDRESS POSTAL CITY HEAD OFFICE CYPRUS Central Banks CY000001 Central Bank of Cyprus 80, Tzon Kennenty Avenue 1076 Nicosia Total number of Central Banks : 1 Credit Institutions CY130001 Allied Bank SAL 276, Archiepiskopou Makariou III Avenue 3105 Limassol LB Allied Bank SAL CY110001 Alpha Bank Limited 1, Prodromou Street 1095 Nicosia CY130002 Arab Bank plc 1, Santaroza Avenue 1075 Nicosia JO Arab Bank plc CY120001 Arab Bank plc 1, Santaroza Avenue 1075 Nicosia JO Arab Bank plc CY130003 Arab Jordan Investment Bank SA 23, Olympion Street 3035 Limassol JO Arab Jordan Investment Bank SA CY130006 Bank of Beirut and the Arab Countries SAL 135, Archiepiskopou Makariou III Avenue 3021 Limassol LB Bank of Beirut and the Arab Countries SAL CY130032 Bank of Beirut SAL 6, Griva Digeni Street 3106 Limassol LB Bank of Beirut SAL CY110002 Bank of Cyprus Ltd 51, Stasinou Street, Strovolos 2002 Nicosia CY130007 Banque Européenne pour le Moyen - Orient SAL 227, Archiepiskopou Makariou III Avenue 3105 Limassol LB Banque Européenne pour le Moyen - Orient SAL CY130009 Banque SBA 8C, Tzon Kennenty Street 3106 Limassol FR Banque SBA CY130010 Barclays Bank plc 88, Digeni Akrita Avenue 1061 Nicosia GB Barclays Bank plc CY130011 BLOM Bank SAL 26, Vyronos Street 3105 Limassol LB BLOM Bank SAL CY130033 BNP Paribas Cyprus Ltd 319, 28 Oktovriou Street 3105 Limassol CY130012 Byblos Bank SAL 1, Archiepiskopou Kyprianou Street 3036 Limassol LB Byblos Bank SAL CY151414 Co-operative Building Society of Civil Servants Ltd 34, Dimostheni Severi Street 1080 Nicosia -
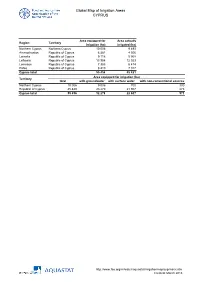
Global Map of Irrigation Areas CYPRUS
Global Map of Irrigation Areas CYPRUS Area equipped for Area actually Region Territory irrigation (ha) irrigated (ha) Northern Cyprus Northern Cyprus 10 006 9 493 Ammochostos Republic of Cyprus 6 581 4 506 Larnaka Republic of Cyprus 9 118 5 908 Lefkosia Republic of Cyprus 13 958 12 023 Lemesos Republic of Cyprus 7 383 6 474 Pafos Republic of Cyprus 8 410 7 017 Cyprus total 55 456 45 421 Area equipped for irrigation (ha) Territory total with groundwater with surface water with non-conventional sources Northern Cyprus 10 006 9 006 700 300 Republic of Cyprus 45 449 23 270 21 907 273 Cyprus total 55 456 32 276 22 607 573 http://www.fao.org/nr/water/aquastat/irrigationmap/cyp/index.stm Created: March 2013 Global Map of Irrigation Areas CYPRUS Area equipped for District / Municipality Region Territory irrigation (ha) Bogaz Girne Main Region Northern Cyprus 82.4 Camlibel Girne Main Region Northern Cyprus 302.3 Girne East Girne Main Region Northern Cyprus 161.1 Girne West Region Girne Main Region Northern Cyprus 457.9 Degirmenlik Lefkosa Main Region Northern Cyprus 133.9 Ercan Lefkosa Main Region Northern Cyprus 98.9 Guzelyurt Lefkosa Main Region Northern Cyprus 6 119.4 Lefke Lefkosa Main Region Northern Cyprus 600.9 Lefkosa Lefkosa Main Region Northern Cyprus 30.1 Akdogan Magosa Main Region Northern Cyprus 307.7 Gecitkale Magosa Main Region Northern Cyprus 71.5 Gonendere Magosa Main Region Northern Cyprus 45.4 Magosa A Magosa Main Region Northern Cyprus 436.0 Magosa B Magosa Main Region Northern Cyprus 52.9 Mehmetcik Magosa Main Region Northern -

Mortuary Landscapes and the Production of Community During the Prehistoric Bronze Age on Cyprus
Dwelling with the Dead: Mortuary Landscapes and the Production of Community During the Prehistoric Bronze Age on Cyprus by Eilis M Monahan This thesis/dissertation document has been electronically approved by the following individuals: Manning,Sturt (Chairperson) Russell,Nerissa (Minor Member) DWELLING WITH THE DEAD: MORTUARY LANDSCAPES AND THE PRODUCTION OF COMMUNITY DURING THE PREHISTORIC BRONZE AGE ON CYPRUS A Thesis Presented to the Faculty of the Graduate School of Cornell University in Partial Fulfillment of the Requirements for the Degree of Master of Arts by Eilis M. Monahan August 2010 © 2010 Eilis M. Monahan ABSTRACT The Prehistoric Bronze Age is a tremendously important period on Cyprus for understanding the later development of the island. During this period architecture, settlement patterns, technologies, and subsistence practices changed drastically, and of greatest interest to this thesis, the dead were buried in discrete cemeteries. It has been historically stated by scholars that these cemeteries were located outside, and within view, of the associated settlements. The recent publication of the excavations and surveys of several Prehistoric Bronze Age sites allows these assumptions to be tested, and their implications considered. The shifts in settlement form and organization and in mortuary behavior clearly reflect major changes in the organization of society during this time period. By analyzing the various relationships between settlement and cemetery within the landscape, this thesis attempts to better understand the dimensions of variation and patterns of practice that shaped the changing society of the Cypriot Bronze Age. Using data from the excavations and surveys at Marki-Alonia, Alambra-Mouttes, and Sotira- Kaminoudhia, and the survey data from the Vasilikos Valley Project, as well as a reconnaissance survey undertaken by the author during the summers of 2008 and 2009, this thesis takes a behavioral and phenomenological approach to the PreBA Cypriot mortuary landscape, addressing four key aspects. -

Name Address Postal City Mfi Id Head Office Res* Cyprus
MFI ID NAME ADDRESS POSTAL CITY HEAD OFFICE RES* CYPRUS Central Banks CY000001 Central Bank of Cyprus 80, Tzon Kennenty Avenue 1076 Nicosia No Total number of Central Banks : 1 Credit Institutions CY110001 Alpha Bank Limited 1, Prodromou Street 1095 Nicosia No CY120001 Arab Bank plc 1, Santaroza Avenue 1075 Nicosia JO Arab Bank plc No CY120004 Arab Jordan Investment Bank SA 23, Olympion Street 3035 Limassol JO Arab Jordan Investment No Bank SA CY120021 Bank of Beirut SAL 6, Griva Digeni Street 3106 Limassol LB Bank of Beirut SAL No CY110002 Bank of Cyprus Public Company Ltd 51, Stasinou Street, Strovolos 2002 Nicosia No CY120026 Bankmed sal 3722 Limassol LB Bankmed sal No CY120006 Banque Européenne pour le Moyen - Orient 227, Archiepiskopou Makariou III 3105 Limassol LB Banque Européenne pour No SAL Avenue le Moyen - Orient SAL CY120007 Banque SBA 8C, Tzon Kennenty Street 3106 Limassol FR Banque SBA No CY120008 Barclays Bank plc 88, Digeni Akrita Avenue 1061 Nicosia GB Barclays Bank plc No CY120005 BBAC SAL 135, Archiepiskopou Makariou III 3021 Limassol LB BBAC SAL No Avenue CY120009 BLOM Bank SAL 26, Vyronos Street 3105 Limassol LB BLOM Bank SAL No CY110018 BNP Paribas Cyprus Ltd 319, 28 Oktovriou Street 3105 Limassol No CY120010 Byblos Bank SAL 1, Archiepiskopou Kyprianou Street 3036 Limassol LB Byblos Bank SAL No CY120023 Central Cooperative Bank PLC 1509 Nicosia BG Central Cooperative Bank No PLC CY151414 Co-operative Building Society of Civil 34, Dimostheni Severi Street 1080 Nicosia No Servants Ltd CY110003 Co-operative Central Bank -

“The Palaepaphos Urban Landscape Project”: Theoretical Background and Preliminary Report 2006-2007
14. (20) p 263_290 c&k 12/4/08 9:54 PM Page 263 “The Palaepaphos Urban Landscape Project”: Theoretical Background and Preliminary Report 2006-2007 Maria Iacovou (University of Cyprus) INTRODUCTION Late Bronze and in the Iron Ages, is to this day poorly understood. Its few visible monuments Fieldwork initiated by the Archaeological —the secular structures in the localities Marchel- Research Unit of the University of Cyprus at Kouklia-Palaepaphos in 2006 has been designed lo, Hadjiabdoulla and Evreti, the sepulchral to meet the requirements of two interconnected projects. The first, to which the present article ____________ provides the theoretical background and explains 1. In my capacity as field director, I wish to thank the Director the 2006 and 2007 field targets, is “The Palaepa- of Antiquities of Cyprus Dr Pavlos Flourentzos for permis- phos Urban Landscape Project”.1 The second, sion to initiate the Palaepaphos-Marchello field project in 2006 and for the annual renewal of his permit. I extend my which is entitled “A long-term response to the gratitude to the Curator of Museums Dr Maria Hadjicosti for need to make modern development and the having encouraged me to consider a long-term excavation preservation of the archaeo-cultural record mutu- project at various key-sites in the vicinity of the sanctuary of ally compatible operations: Pilot application at Palaepaphos and for her sound advice on formal procedures. I thank most warmly the Curator of Monuments Dr Marina Kouklia-Palaepaphos” (“Palaepaphos Pilot Pro- Ieronymidou for her steadfast support of the different aspects ject”, for short), is a project concerned with the of our field operations (geophysical surveys and excavations) development of a framework of principles for the and her impeccable and timely handling of issues as sensitive as the declaration or expropriation of private land. -
S CHOOL BOARDS NICOSIA ΕΠΑΡΧΙΑ: School Board Name
SCHOOL BOARDS ΕΠΑΡΧΙΑ: NICOSIA School Board Name Address Postal Code District Tel Tel Fax Email EFOREIA ARMENIKON SCHOLEION KYPROU P.O.Box 16173 2086 Lefkosia NICOSIA 22422218 22311483 [email protected] SCHOLIKI EFOREIA AGIAS VARVARAS 53 Leof. Grigori Afxentiou 2560 Agia Varvara NICOSIA 22522270 22523711 [email protected] SCHOLIKI EFOREIA AGIAS MARINAS XYLIATOU 2772 Agia Marina Xyliatou NICOSIA 22852775 22852847 [email protected] SCHOLIKI EFOREIA AGIOU DOMETIOU 23 Kentavrou 2364 Agios Dometios NICOSIA 22777759 22776570 [email protected] SCHOLIKI EFOREIA AGIOU EPIFANIOU 2610 Agios Epifanios NICOSIA 22633901 22633902 [email protected] SCHOLIKI EFOREIA AGIOU IOANNI LEFKOSIAS Apostolou Andrea 1 2611 Agios Ioannis Lefkosias NICOSIA 22634311 22634310 [email protected] SCHOLIKI EFOREIA AGIOU MARONA 20 Agiou Marona 2304 Anthoupoli NICOSIA 22721510 22372205 [email protected] SCHOLIKI EFOREIA AGION TRIMITHIAS 40 Archiepiskopou Makariou III 2671 Agioi Trimithias NICOSIA 22832470 22835054 [email protected] SCHOLIKI EFOREIA AGLANTZIAS P.O.Box 20266 2150 Aglantzia NICOSIA 22333847 22337901 [email protected] SCHOLIKI EFOREIA AGROKIPIAS 2617 Agrokipia NICOSIA 22634111 22634111 [email protected] SCHOLIKI EFOREIA AKAKIOU Grigori Afxentiou 2720 Akaki NICOSIA 22823466 22824524 [email protected] SCHOLIKI EFOREIA ALAMPRAS 2563 Alampra NICOSIA 22522457 22526402 [email protected] SCHOLIKI EFOREIA -

Authentic Route 10
Cyprus Authentic Route 10 Safety Driving in Cyprus Comfort Rural Accommodation Tips Useful Information Only DIGITAL Version Ecclesiastical Treasures Lefkosia • Strovolos • Kato and Pano Deftera • Psimolofou • Episkopio • Politiko-Tamassos • Pera • Kampia • Kapedes • Machairas Monastery • Lazanias • Gourri • Fikardou • Klirou • Malounta • Agrokipia • Mitsero • Platanistasa • Fterikoudi • Askas • Palaichori • Alona • Polystypos • Lagoudera • Xyliatos • Agia Marina • Orounta • Peristerona • Akaki • Kokkinotrimithia • Lefkosia Route 10 Lefkosia – Strovolos – Kato and Pano Deftera – Psimolofou – Episkopio – Politiko-Tamassos – Pera – Kampia – Kapedes – Machairas Monastery – Lazanias – Gourri – Fikardou – Klirou – Malounta – Agrokipia – Mitsero – Platanistasa – Fterikoudi – Askas – Palaichori – Alona – Polystypos – Lagoudera – Xyliatos – Agia Marina – Orounta – Peristerona – Akaki – Kokkinotrimithia – Lefkosia scale 1:350,000 Skylloura Agia Kanli Mandres Kioneli 0 1 2 4 6 8 Kilometers Marina Agios Kyra Vasileios Morfou Mia Ortakioi Milia Masari Argaki Fyllia Gerolakkos Trachonas Kato Mammari Agios Katokopia Deneia Nikitas Zodeia Dometios Pano Nicosia International Egkomi Aglantzia Zodeia Avlona Kokkinotrimithia Airport (Not in use) Astromeritis Akaki Strovolos Peristerona Agioi Angolemi Palaiometocho Lakatameia Trimithias Geri Potami Orounta Meniko Anthoupoli Latsia Pano Kato Koutrafas Pano Panagia Koutrafas Deftera Chrysospiliotissa Nikitari Agios Kato Kato Agios Deftera Vyzakia Agioi Ioannis Anageia Tseri Moni Panteleimon Vyzakia Iliofotoi -

Pagemaker Econ Stat 2003 300805
ȀȊȆȇǿǹȀǾ ǻǾȂȅȀȇǹȉǿǹ REPUBLIC OF CYPRUS ȈȉǹȉǿȈȉǿȀȅǿ ȀȍǻǿȀȅǿ ǻǾȂȍȃ, ȀȅǿȃȅȉǾȉȍȃ Ȁǹǿ Ǽȃȅȇǿȍȃ ȉǾȈ ȀȊȆȇȅȊ STATISTICAL CODES OF MUNICIPALITIES, COMMUNITIES AND QUARTERS OF CYPRUS 2010 ȈȉǹȉǿȈȉǿȀǾ ȊȆǾȇǼȈǿǹ STATISTICAL SERVICE ȈIJĮIJȚıIJȚțȠȓ ȀȦįȚțȠȓ: x ȈİȚȡȐ ǿ x ǹȡ. DzțșİıȘȢ 6 Statistical Codes: x ȈİȚȡȐ ǿ x Report No 6 - 3 - ȆȇȅȁȅīȅȈ PREFACE ǹȣIJȒ İȓȞĮȚ Ș ȑțIJȘ ȑțįȠıȘ IJȠȣ įȘµȠıȚİȪµĮIJȠȢ This is the sixth edition of the publication “Statistical «ȈIJĮIJȚıIJȚțȠȓ ȀȦįȚțȠȓ ǻȒµȦȞ, ȀȠȚȞȠIJȒIJȦȞ țĮȚ ǼȞȠȡȚȫȞ Codes of Municipalities, Communities and Quarters of IJȘȢ ȀȪʌȡȠȣ» țĮȚ ʌĮȡȠȣıȚȐȗİȚ IJȠ ĮȞĮșİȦȡȘµȑȞȠ Cyprus” and it presents the updated statistical coding ıIJĮIJȚıIJȚțȩ țȦįȚțȩ ıȪıIJȘµĮ IJȦȞ IJȠʌȚțȫȞ İįĮijȚțȫȞ system of the local territorial units (municipalities, µȠȞȐįȦȞ (įȒµȦȞ, țȠȚȞȠIJȒIJȦȞ țĮȚ İȞȠȡȚȫȞ) IJȘȢ ȀȪʌȡȠȣ. communities and quarters) of Cyprus. The revised ȉȠ ĮȞĮșİȦȡȘµȑȞȠ ıȪıIJȘµĮ ȖİȦȖȡĮijȚțȫȞ țȦįȚțȫȞ geographical coding system was prepared in ȑȖȚȞİ ıİ ıȣȞİȡȖĮıȓĮ µİ ȐȜȜİȢ ĮȡµȩįȚİȢ ȊʌȘȡİıȓİȢ țĮȚ collaboration with other competent Services and ȉµȒµĮIJĮ țĮȚ ıIJȠȤİȪİȚ ıIJȘȞ İȣȡȪIJİȡȘ įȣȞĮIJȒ ȤȡȒıȘ. Departments and aims at its broadest possible use. Ǿ ȖİȦȖȡĮijȚțȒ IJĮȟȚȞȩµȘıȘ ʌİȡȚȜĮµȕȐȞİȚ IJȚȢ įȚȠȚțȘIJȚțȑȢ The geographical classification pertains to the İʌĮȡȤȓİȢ, IJȠȣȢ įȒµȠȣȢ, IJȚȢ țȠȚȞȩIJȘIJİȢ țĮȚ IJȚȢ İȞȠȡȓİȢ. administrative districts, the municipalities, the ȉĮ ȠȞȩµĮIJĮ IJȦȞ įȒµȦȞ/țȠȚȞȠIJȒIJȦȞ țĮȚ İȞȠȡȚȫȞ İȓȞĮȚ IJĮ communities and the quarters. The names of İʌȓıȘµĮ ȠȞȩµĮIJĮ µİ IJȘ ȖȡĮijȒ ȩʌȦȢ țĮșȠȡȓıIJȘțİ Įʌȩ municipalities/communities and quarters are the official IJȘ ȂȩȞȚµȘ ȀȣʌȡȚĮțȒ ǼʌȚIJȡȠʌȒ ȉȣʌȠʌȠȓȘıȘȢ names which have been determined by the Cyprus īİȦȖȡĮijȚțȫȞ ȅȞȠµȐIJȦȞ ıIJȘȞ ǼȜȜȘȞȚțȒ ȖȜȫııĮ țĮȚ IJȘ Permanent Committee for the Standardization of µİIJĮȖȡĮijȒ IJȠȣȢ ıIJȠ ȇȦµĮȞȚțȩ ĮȜijȐȕȘIJȠ. Geographical Names in the Greek language and transliterated in Roman characters. ĬĮ ʌȡȑʌİȚ ȞĮ ıȘµİȚȦșİȓ ȩIJȚ IJȠ țȦįȚțȩ ıȪıIJȘµĮ It should be noted that the coding system covers the țĮȜȪʌIJİȚ ȠȜȩțȜȘȡȘ IJȘȞ ȀȪʌȡȠ, įȘȜ. -

Cypriot Material Culture Studies from Picrolite Carving to Proskynitaria Analysis
POCA 2008 Ariane JACOBS & Peter COSYNS (eds.) CYPRIOT MATERIAL CULTURE STUDIES FROM PICROLITE CARVING TO PROSKYNITARIA ANALYSIS Proceedings of the 8th Annual Postgraduate Cypriot Archaeology Conference Held in Honour of the Memory of Paul Åström at the Vrije Universiteit Brussel (Belgium), 27th – 29th November 2008 © 2015 Uitgeverij VUBPRESS Brussels University Press VUBPRESS is an imprint of ASP nv (Academic & Scientific Publishers nv) Keizerslaan 34 B-1000 Brussels Tel. +32 (0)2 289 26 50 Fax +32 (0)2 289 26 59 E-mail: [email protected] www.vubpress.be ISBN 978 90 5487 974 9 NUR 654 Legal deposit D/2015/11.161/097 All rights reserved. No parts of this book may be reproduced or transmitted in any form or by any means, electronic, mechanical, photocopying, recording, or otherwise, without the prior written permission of the author. II Table of contents Bibliographic Abbreviations Acknowledgements Ariane JACOBS & Peter Preface COSYNS Jean Paul VAN BENDEGEM Foreword Karin NYS Paul Åström, The Mentor of Several Generations of Archaeological Students and an Inspiration for Generations to Come Paul ÅSTRÖM† Cypriot Archaeology: a Prospering History Priscilla KESWANI Olive Production, Storage, and Political Economy at Late 1 Bronze Age Kalavasos, Cyprus Elizabeth CORY-LOPEZ Technological and Material Approaches to Cypriot Middle 25 Chalcolithic Picrolite Michelle GAMBLE & Kirsi Assessing Growth Disruptions and Preservation Levels: 47 LORENTZ Preliminary Results on Linear Enamel Hypoplasias at the Souskiou-Laona Chalcolithic Cemetery, Cyprus Jan -

Name Outlet Address Location Phone 16 DEKAEXI ONASAGOROU 16
Name Outlet Address Location Phone 16 DEKAEXI ONASAGOROU 16, STOA KLOKKARI NICOSIA 99377297, 2nd line:22662227 20-4-7 YPERAGORA GREGORIOU ANAGNOSTOPOULOU 12 NICOSIA 99221854, 2nd line:22324422 34 THE SHOP ARCH MAKARIOU AVE 17 NICOSIA 22758428 371 BOUTIQUE ARCH MAKARIOU III 25, SHOP 2 NICOSIA 99275247 48 BISTRO MAKARIOS AVE 48 NICOSIA 99680441 5 A SEC DRY CLEANING IFIGENIAS 46, SHOP H-Z NICOSIA 22261631, 2nd line:99340478 7 STARS PIZZA PERIKLEOUS 13, SHOP B-C NICOSIA 99485303 96SPORTSMANIA ARCH. MAKARIOU III 27C NICOSIA 22525390, 2nd line:99547499 A & A OPTICAL CENTER HOMER AVE.29A NICOSIA 22664853, 2nd line:99646775 A MAVRIS & SONS LTD (EPIP ERODOTOU 13, GERI NICOSIA 22430384, 2nd line:99442992 A&A ANTONIOU PHARMACIES L LEOF. GERIOU 26 NICOSIA 22487810, 2nd line:99963081 A&A DOROS PERFOMANCE TUNI KYRIAKOU MATSI 98 NICOSIA 22591212, 2nd line:99406106 A&X ANTONIOU ARSINOES 11C NICOSIA 22490777 A. KRONFOUL TRADING LTD DIOS 4C, LATSIA INDUSTRIAL AREA NICOSIA 22489084, 2nd line:99888069 A. KRONFOUL TRADING LTD GONIA EVRIPIDOU & ONASAGOROU NICOSIA 22681894 A. KRONFOUL TRADING LTD GREGORI AFXENTIOU 65 A-Z NICOSIA 22344032 A. KXOFFEE PROJECT PINDAROU 29D NICOSIA 99559776, 2nd line:22831324 A. TSIAPINIS YPERAGORA MAKARIOU 16 NICOSIA 22832392, 2nd line:99683675 A.A. SPORTRACING LTD ARCHANGELOU MICHAEL 40B NICOSIA 22872838, 2nd line:99420067 A.A.PAZZOLI LTD 28TH OCTOBER STREET 56D NICOSIA 22358004, 2nd line:22358004 A.C. NICOLAOU BLINDS LTD ATHALASSIS AVENUE 76A NICOSIA 22513139, 2nd line:22621119 A.C.I. CAR SCAN SERVICES AYIOU ILARIONOS 24D, KAIMAKLI NICOSIA 22430100, 2nd line:99676991 A.CH. BEAUTY & FITNESS LT EVANTHOUS 24 NICOSIA 22459900, 2nd line:24819525 A.I.MOTOKINISI LTD SERVIC MEGARON 15 NICOSIA 22353994, 2nd line:77771911 A.KANAKA AKTINOLOGIKO KYRIAKOU MATSI AVE 11, NIKIS CENTRE 2ND FLOOR NICOSIA 22318600 A.M.P.M. -

Map 72 Cyprus Compiled by D
Map 72 Cyprus Compiled by D. Rupp, 1994 Introduction Many factors, both ancient and not so ancient, have contributed to the cultural map that can be constructed of Cyprus between 1100 B.C. and A.D. 640. Before the beginning of the third century B.C. the island remained on the periphery of the Greek world. Only then did hellenization begin to permeate its entire society, not just the elite in the urban centers of some of the Iron Age kingdoms. Political and economic ties with the Levant, as well as cultural influences, had contributed significantly to shaping the Cypriote landscape in the Iron Age. Until hellenization irrevocably transformed Cypriote society and its use of the landscape in the Hellenistic period, the historical geography of the island must be understood in its own indigenous terms. Aside from the principal urban centers of the Iron Age kingdoms, Greek and Roman sources do not provide a wealth of detailed information on the geography of Cyprus. Extant epigraphical sources are likewise relatively mute on this topic. It is thus archaeology that must be employed as the primary source for historical geography. In the later nineteenth century, the plundering of chamber tombs and of rural sanctuaries under the guise of research was the norm in Cypriote archaeology. Although subsequently this regrettable approach was abandoned and the scientific excavation of kingdom centers commenced, fascination with the material culture revealed by tombs and by sanctuaries continued unabated. As a result, while there now exists a wealth of evidence for their location, overall distribution patterns and periods of occupation, by contrast the location, size and nature of settlements is much less well understood.