The Contribution of Fossils to the Reconstruction of the Higher- Level Phylogeny of Birds
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

6-A John James Audubon, American Flamingo, 1838
JOHN JAMES AUDUBON [1785–1851] 6 a American Flamingo,1838 American Flamingo is one of the 435 hand-colored engravings that River, a major flyway for migratory birds, and eventually wan- make up John James Audubon’s monumental Birds of America, dered farther from home to comb the American frontier for issued in four volumes between 1826 and 1838. The massive unrecorded species. publication includes life-size representations of nearly five hundred Audubon’s procedure was to study and sketch a bird in its natural species of North American birds. Although Audubon was not the habitat before killing it carefully, using fine shot to minimize dam- first to attempt such a comprehensive catalog, his work departed age. His critical innovation was to then thread wire through the from conventional scientific illustration, which showed lifeless spec- specimen, allowing him to fashion a lifelike pose. He worked in imens against a blank background, by presenting the birds as they watercolor, and had completed some four hundred paintings appeared in the wild. When his pictures were first published, when he decided to publish them as a folio of prints. Failing to find some naturalists objected to Audubon’s use of dramatic action and support in Philadelphia, he sailed for England, where he became pictorial design, but these are the qualities that set his work apart lionized as “The American Woodsman.” The engraving firm and make it not only an invaluable record of early American Robert Havell and Son took on the challenge of reproducing wildlife but an unmatched work of American art. Audubon’s paintings on copper plates and tinting the resulting John James Audubon was born in Haiti and educated in France, black-and-white prints by hand. -

A Pliocene Flamingo from Mexico
June, 1944 THE WILSON BULLETIN 77 Vol. 56. No. 2 A PLIOCENE FLAMINGO FROM MEXICO BY LOYE MILLER IELD parties from the California Institute of Technology have been F fortunate in locating a variety of fossil deposits in Mexico that in- cluded bird remains. Some have been very rich in the quantity and variety of material; for example, the San Josecito Cavern of Nuevo Leon (Miller, ,1943), a deposit of Pleistocene age, yielded several thousand bird bones assigned to over forty species. The present paper deals with a collection of ten fragments, all but one of which are in- cluded in a single species. I am indebted to Dr. Chester Stock in charge of the explorations for the opportunity of working with the bird collec- tions. Dr. Alexander Wetmore has loaned comparative material, and Dr. Hildegarde Howard has been a most congenial fellow student during many conferences on the flamingoes, both Recent and Fossil. To these several colleagues my sincere thanks are offered. The ten fragments are from collecting locality No. 289, California Institute of Technology, known. as the Rincon Pliocene, Chihuahua, Mexico. Associated mammal remains include horse, camel, antelope, and carnivore species. The matrix is a fine grained silt of lightest color, without cementing material. A stiff brush serves to remove it from the well petrified bones. Unfortunately the specimens are most frag- mentary. They do, however, prove to be of interest in several respects; most notably they prove (since several speciments are from pre-volant young) that a small speciesof flamingo was present as a breeding bird. This is the earliest record for the family in America. -

Flamingo ABOUT the GROUP
Flamingo ABOUT THE GROUP Bulletin of the IUCN-SSC/Wetlands International The Flamingo Specialist Group (FSG) was established in 1978 at Tour du Valat in France, under the leadership of Dr. Alan Johnson, who coordinated the group until 2004 (see profile at www.wetlands.org/networks/Profiles/January.htm). Currently, the group is FLAMINGO SPECIALIST GROUP coordinated from the Wildfowl & Wetlands Trust at Slimbridge, UK, as part of the IUCN- SSC/Wetlands International Waterbird Network. The FSG is a global network of flamingo specialists (both scientists and non- scientists) concerned with the study, monitoring, management and conservation of the world’s six flamingo species populations. Its role is to actively promote flamingo research and conservation worldwide by encouraging information exchange and cooperation amongst these specialists, and with other relevant organisations, particularly IUCN - SSC, Ramsar, WWF International and BirdLife International. FSG members include experts in both in-situ (wild) and ex-situ (captive) flamingo conservation, as well as in fields ranging from field surveys to breeding biology, diseases, tracking movements and data management. There are currently 165 members around the world, from India to Chile, and from France to South Africa. Further information about the FSG, its membership, the membership list serve, or this bulletin can be obtained from Brooks Childress at the address below. Chair Assistant Chair Dr. Brooks Childress Mr. Nigel Jarrett Wildfowl & Wetlands Trust Wildfowl & Wetlands Trust Slimbridge Slimbridge Glos. GL2 7BT, UK Glos. GL2 7BT, UK Tel: +44 (0)1453 860437 Tel: +44 (0)1453 891177 Fax: +44 (0)1453 860437 Fax: +44 (0)1453 890827 [email protected] [email protected] Eastern Hemisphere Chair Western Hemisphere Chair Dr. -
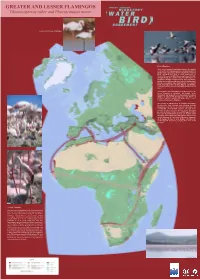
GREATER and LESSER FLAMINGOS Phoenicopterus Ruber and Phoeniconaias Minor
GREATER AND LESSER FLAMINGOS Phoenicopterus ruber and Phoeniconaias minor Greater and Lesser Flamingos © Cliff Buckton © P & H Harris Lesser Flamingo The Lesser Flamingo Phoeniconaias minor is the smallest of the world's five flamingo species. It occurs primarily in the Rift Valley lakes of East Africa with about 4 to 5 million birds estimated, but also in small populations in Namibia/Botswana (40,000), Mauritania/Senegal (15,400), Ethiopia (8,300). The alkaline lakes of the Rift Valley are the primary feeding areas for the East Africa population. During non-breeding periods these lakes often hold almost the entire population. Huge feeding flocks of 1-2 million birds frequently gather on lakes Bogoria and Nakuru, creating one of the most stunning wildlife spectacles in the world. Although it is still the most numerous of the five species, the Lesser Flamingo is classified as globally "near threatened" due primarily to its dependence on a limited number of unprotected breeding sites and threats of proposed soda-ash mining and hydro-electric power schemes on the main breeding lakes. The question of whether there is occasional interchange between the East African and southern African populations has yet to be resolved definitely, but considerable circumstantial evidence has now been assembled to show that East African Lesser Flamingos probably do fly to Botswana to breed during periods when the Lake Makgadikgadi Salt Pans are flooded. Their migration routes, flight range and stopover places (if any) are still unknown. It is now known that Lesser Flamingos do fly during the day, at great heights, well above the normal diurnal movement of eagles, their main aerial predator. -

Flamingo Newsletter 17, 2009
ABOUT THE GROUP The Flamingo Specialist Group (FSG) is a global network of flamingo specialists (both scientists and non-scientists) concerned with the study, monitoring, management and conservation of the world’s six flamingo species populations. Its role is to actively promote flamingo research, conservation and education worldwide by encouraging information exchange and cooperation among these specialists, and with other relevant organisations, particularly the IUCN Species Survival Commission (SSC), the Ramsar Convention on Wetlands, the Convention on Conservation of Migratory Species (CMS), the African-Eurasian Migratory Waterbird Agreement (AEWA), and BirdLife International. The group is coordinated from the Wildfowl & Wetlands Trust, Slimbridge, UK, as part of the IUCN-SSC/Wetlands International Waterbird Network. FSG members include experts in both in-situ (wild) and ex-situ (captive) flamingo conservation, as well as in fields ranging from research surveys to breeding biology, infectious diseases, toxicology, movement tracking and data management. There are currently 286 members representing 206 organisations around the world, from India to Chile, and from France to South Africa. Further information about the FSG, its membership, the membership list serve, or this bulletin can be obtained from Brooks Childress at the address below. Chair Dr. Brooks Childress Wildfowl & Wetlands Trust Slimbridge Glos. GL2 7BT, UK Tel: +44 (0)1453 860437 Fax: +44 (0)1453 860437 [email protected] Eastern Hemisphere Chair Western Hemisphere Chair Dr. Arnaud Béchet Dr. Felicity Arengo Station biologique, Tour du Valat American Museum of Natural History Le Sambuc Central Park West at 79th Street 13200 Arles, France New York, NY 10024 USA Tel : +33 (0) 4 90 97 20 13 Tel: +1 212 313-7076 Fax : +33 (0) 4 90 97 20 19 Fax: +1 212 769-5292 [email protected] [email protected] Citation: Childress, B., Arengo, F. -

Onetouch 4.0 Scanned Documents
/ Chapter 2 THE FOSSIL RECORD OF BIRDS Storrs L. Olson Department of Vertebrate Zoology National Museum of Natural History Smithsonian Institution Washington, DC. I. Introduction 80 II. Archaeopteryx 85 III. Early Cretaceous Birds 87 IV. Hesperornithiformes 89 V. Ichthyornithiformes 91 VI. Other Mesozojc Birds 92 VII. Paleognathous Birds 96 A. The Problem of the Origins of Paleognathous Birds 96 B. The Fossil Record of Paleognathous Birds 104 VIII. The "Basal" Land Bird Assemblage 107 A. Opisthocomidae 109 B. Musophagidae 109 C. Cuculidae HO D. Falconidae HI E. Sagittariidae 112 F. Accipitridae 112 G. Pandionidae 114 H. Galliformes 114 1. Family Incertae Sedis Turnicidae 119 J. Columbiformes 119 K. Psittaciforines 120 L. Family Incertae Sedis Zygodactylidae 121 IX. The "Higher" Land Bird Assemblage 122 A. Coliiformes 124 B. Coraciiformes (Including Trogonidae and Galbulae) 124 C. Strigiformes 129 D. Caprimulgiformes 132 E. Apodiformes 134 F. Family Incertae Sedis Trochilidae 135 G. Order Incertae Sedis Bucerotiformes (Including Upupae) 136 H. Piciformes 138 I. Passeriformes 139 X. The Water Bird Assemblage 141 A. Gruiformes 142 B. Family Incertae Sedis Ardeidae 165 79 Avian Biology, Vol. Vlll ISBN 0-12-249408-3 80 STORES L. OLSON C. Family Incertae Sedis Podicipedidae 168 D. Charadriiformes 169 E. Anseriformes 186 F. Ciconiiformes 188 G. Pelecaniformes 192 H. Procellariiformes 208 I. Gaviiformes 212 J. Sphenisciformes 217 XI. Conclusion 217 References 218 I. Introduction Avian paleontology has long been a poor stepsister to its mammalian counterpart, a fact that may be attributed in some measure to an insufRcien- cy of qualified workers and to the absence in birds of heterodont teeth, on which the greater proportion of the fossil record of mammals is founded. -

1 ID Euring Latin Binomial English Name Phenology Galliformes
BIRDS OF METAURO RIVER: A GREAT ORNITHOLOGICAL DIVERSITY IN A SMALL ITALIAN URBANIZING BIOTOPE, REQUIRING GREATER PROTECTION 1 SUPPORTING INFORMATION / APPENDICE Check list of the birds of Metauro river (mouth and lower course / Fano, PU), up to September 2020. Lista completa delle specie ornitiche del fiume Metauro (foce e basso corso /Fano, PU), aggiornata ad Settembre 2020. (*) In the study area 1 breeding attempt know in 1985, but in particolar conditions (Pandolfi & Giacchini, 1985; Poggiani & Dionisi, 1988a, 1988b, 2019). ID Euring Latin binomial English name Phenology GALLIFORMES Phasianidae 1 03700 Coturnix coturnix Common Quail Mr, B 2 03940 Phasianus colchicus Common Pheasant SB (R) ANSERIFORMES Anatidae 3 01690 Branta ruficollis The Red-breasted Goose A-1 (2012) 4 01610 Anser anser Greylag Goose Mi, Wi 5 01570 Anser fabalis Tundra/Taiga Bean Goose Mi, Wi 6 01590 Anser albifrons Greater White-fronted Goose A – 4 (1986, february and march 2012, 2017) 7 01520 Cygnus olor Mute Swan Mi 8 01540 Cygnus cygnus Whooper Swan A-1 (1984) 9 01730 Tadorna tadorna Common Shelduck Mr, Wi 10 01910 Spatula querquedula Garganey Mr (*) 11 01940 Spatula clypeata Northern Shoveler Mr, Wi 12 01820 Mareca strepera Gadwall Mr, Wi 13 01790 Mareca penelope Eurasian Wigeon Mr, Wi 14 01860 Anas platyrhynchos Mallard SB, Mr, W (R) 15 01890 Anas acuta Northern Pintail Mi, Wi 16 01840 Anas crecca Eurasian Teal Mr, W 17 01960 Netta rufina Red-crested Pochard A-4 (1977, 1994, 1996, 1997) 18 01980 Aythya ferina Common Pochard Mr, W 19 02020 Aythya nyroca Ferruginous -

A Skull of a Very Large Crane from the Late Miocene of Southern Germany, with Notes on the Phylogenetic Interrelationships of Extant Gruinae
Journal of Ornithology https://doi.org/10.1007/s10336-020-01799-0 ORIGINAL ARTICLE A skull of a very large crane from the late Miocene of Southern Germany, with notes on the phylogenetic interrelationships of extant Gruinae Gerald Mayr1 · Thomas Lechner2 · Madelaine Böhme2 Received: 3 June 2020 / Revised: 1 July 2020 / Accepted: 2 July 2020 © The Author(s) 2020 Abstract We describe a partial skull of a very large crane from the early late Miocene (Tortonian) hominid locality Hammerschmiede in southern Germany, which is the oldest fossil record of the Gruinae (true cranes). The fossil exhibits an unusual preservation in that only the dorsal portions of the neurocranium and beak are preserved. Even though it is, therefore, very fragmentary, two morphological characteristics are striking and of paleobiological signifcance: its large size and the very long beak. The fossil is from a species the size of the largest extant cranes and represents the earliest record of a large-sized crane in Europe. Overall, the specimen resembles the skull of the extant, very long-beaked Siberian Crane, Leucogeranus leucogeranus, but its afnities within Gruinae cannot be determined owing to the incomplete preservation. Judging from its size, the fossil may possibly belong to the very large “Grus” pentelici, which stems from temporally and geographically proximate sites. The long beak of the Hammerschmiede crane conforms to an open freshwater paleohabitat, which prevailed at the locality. Keywords Aves · Evolution · Gruidae · Systematics · Tortonian Zusammenfassung Ein Schädel eines sehr großen Kranichs aus dem Obermiozän Süddeutschlands, mit Bemerkungen zu den Verwandtschaftsbeziehungen heutiger Gruinae Wir beschreiben einen partiell erhaltenen Schädel eines sehr großen Kranichs aus dem frühen Obermiozän (Tortonium) der Hominiden-Fundstelle Hammerschmiede in Süddeutschland, welcher den ältesten Fossilnachweis der Gruinae (echte Kraniche) repräsentiert. -

AOU Classification Committee – North and Middle America
AOU Classification Committee – North and Middle America Proposal Set 2016-C No. Page Title 01 02 Change the English name of Alauda arvensis to Eurasian Skylark 02 06 Recognize Lilian’s Meadowlark Sturnella lilianae as a separate species from S. magna 03 20 Change the English name of Euplectes franciscanus to Northern Red Bishop 04 25 Transfer Sandhill Crane Grus canadensis to Antigone 05 29 Add Rufous-necked Wood-Rail Aramides axillaris to the U.S. list 06 31 Revise our higher-level linear sequence as follows: (a) Move Strigiformes to precede Trogoniformes; (b) Move Accipitriformes to precede Strigiformes; (c) Move Gaviiformes to precede Procellariiformes; (d) Move Eurypygiformes and Phaethontiformes to precede Gaviiformes; (e) Reverse the linear sequence of Podicipediformes and Phoenicopteriformes; (f) Move Pterocliformes and Columbiformes to follow Podicipediformes; (g) Move Cuculiformes, Caprimulgiformes, and Apodiformes to follow Columbiformes; and (h) Move Charadriiformes and Gruiformes to precede Eurypygiformes 07 45 Transfer Neocrex to Mustelirallus 08 48 (a) Split Ardenna from Puffinus, and (b) Revise the linear sequence of species of Ardenna 09 51 Separate Cathartiformes from Accipitriformes 10 58 Recognize Colibri cyanotus as a separate species from C. thalassinus 11 61 Change the English name “Brush-Finch” to “Brushfinch” 12 62 Change the English name of Ramphastos ambiguus 13 63 Split Plain Wren Cantorchilus modestus into three species 14 71 Recognize the genus Cercomacroides (Thamnophilidae) 15 74 Split Oceanodroma cheimomnestes and O. socorroensis from Leach’s Storm- Petrel O. leucorhoa 2016-C-1 N&MA Classification Committee p. 453 Change the English name of Alauda arvensis to Eurasian Skylark There are a dizzying number of larks (Alaudidae) worldwide and a first-time visitor to Africa or Mongolia might confront 10 or more species across several genera. -

NEWS and NOTES by Paul Hess
NEWS AND NOTES by Paul Hess First-year Migrants sumably innate southwesterly bearing, and many ended up May Need Experience in Iberia far south of the usual winter range. His report in 1958 suggests that adults improve their inherent naviga - Inheritance or experience? Genetic programming or practi - tional map based on experience, but that first-year birds’ di - cal learning? Various studies show that both inherited and rectional behavior has not progressed beyond an inherited learned information helps birds to migrate to the right rudimentary compass ( Ardea 46:1–37). place in fall and to return to the right place in spring. Im - Fifty years later, Kasper Thorup and six coauthors have mature birds apparently use an innate compass that may added a radio-tracking dimension to similar research using not always allow them to deviate from “hard-wired” direc - White-crowned Sparrows. Results they report in 2007 are a near-perfect analogy to Perdeck’s findings, except that long- distance, non-social, nocturnal migrants are studied rather than short-distance, social, diurnal migrants ( Proceedings of the National Academy of Sciences 104:18115–18119). Fif - teen adult and 15 first-year southbound migrants of the gambelii subspecies were captured at a fall stopover site in Washington state and transported to New Jersey, where each was fitted with a tiny radio transmitter. The birds’ ini - tial movements near the release site were monitored by ground crews, but after flying away from the immediate area (usually at night) they were tracked individually by aircraft for distances as far as 122 kilometers. Adults and immatures rapidly demonstrated a conspicu - ous behavioral difference. -

Fossil Birds of the Nebraska Region
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Transactions of the Nebraska Academy of Sciences and Affiliated Societies Nebraska Academy of Sciences 1992 Fossil Birds of the Nebraska Region James E. Ducey [email protected] Follow this and additional works at: https://digitalcommons.unl.edu/tnas Part of the Life Sciences Commons Ducey, James E., "Fossil Birds of the Nebraska Region" (1992). Transactions of the Nebraska Academy of Sciences and Affiliated Societies. 130. https://digitalcommons.unl.edu/tnas/130 This Article is brought to you for free and open access by the Nebraska Academy of Sciences at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in Transactions of the Nebraska Academy of Sciences and Affiliated Societiesy b an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. 1992. Transactions of the Nebraska Academy of Sciences, XIX: 83-96 FOSSIL BIRDS OF THE NEBRASKA REGION James Ducey 235 Nebraska Hall Lincoln, Nebraska 68588-0521 ABSTRACT Crane (Grus haydeni = Grus canadensis) (Marsh, 1870) and a species of hawk (Buteo dananus) from along the This review compiles published and a few unpublished Loup Fork (Marsh, 1871). records offossil and prehistoric birds for the Nebraska region (Nebraska and parts of adjacent states) from the Cretaceous Many ofthe species first described were from mate Period to the late Pleistocene, about 12,000 years before present. Species recorded during the various epochs include: rial collected in the Great Plains region, including Kan Oligocene and Early Miocene (13 families; 29 species), Middle sas and Wyoming (Marsh, 1872b). The work of scien Miocene (six families; 12 species), Late Miocene (14 families; tists associated with the University of Nebraska in 21 species), Pliocene (six families; 15 species), Early-Middle cluded studies made around the turn-of-the-century. -

Columbiformes ~ Pterocliformes ~ Mesitornithiformes
Birds of the World part 2 Galloanseres, Neoaves: Columbea NEOGNATHAE (the rest of the birds!): Galloanseres • ORDER ANSERIFORMES – waterfowl • Family Anhimidae – screamers (3 species) • Family Anseranatidae – magpie goose (1 species) • Family Anatidae – ducks, geese, and swans (173 species) • ORDER GALLIFORMES – landfowl • Family Megapodiidae – megapodes (21 species) • Family Cracidae – chachalacas, curassows, and guans (55 species) • Family Numididae – guineafowl (6 species) • Family Odontophoridae – New World quail (34 species) • Family Phasianidae – pheasants and allies (183 species) NEOGNATHAE : Neoaves (the rest of the birds!): COLUMBEA • ORDER PODICIPEDIFORMES – Family Podicipedidae – grebes (23 species) • ORDER PHOENICOPTERIFORMES – Family Phoenicopteridae – flamingos (6 species) • ORDER COLUMBIFORMES – Family Columbidae – pigeons and doves (334 species) • ORDER PTEROCLIDIFORMES – Family Pteroclididae – sandgrouse (16 species) • ORDER MESITORNITHIFORMES – Family Mesitornithidae – mesites (3 species) NEOGNATHAE : Galloanseres • ORDER ANSERIFORMES – waterfowl • Family Anhimidae – screamers (3 species) • Family Anseranatidae – magpie goose (1 species) • Family Anatidae – ducks, geese, and swans (173 species) • ORDER GALLIFORMES – landfowl • Family Megapodiidae – megapodes (21 species) • Family Cracidae – chachalacas, curassows, and guans (55 species) • Family Numididae – guineafowl (6 species) • Family Odontophoridae – New World quail (34 species) • Family Phasianidae – pheasants and allies (183 species) southern or crested screamer