The Amarna South Tombs Cemetery
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Origins and Consequences of Kin Networks and Marriage Practices
The origins and consequences of kin networks and marriage practices by Duman Bahramirad M.Sc., University of Tehran, 2007 B.Sc., University of Tehran, 2005 Thesis Submitted in Partial Fulfillment of the Requirements for the Degree of Doctor of Philosophy in the Department of Economics Faculty of Arts and Social Sciences c Duman Bahramirad 2018 SIMON FRASER UNIVERSITY Summer 2018 Copyright in this work rests with the author. Please ensure that any reproduction or re-use is done in accordance with the relevant national copyright legislation. Approval Name: Duman Bahramirad Degree: Doctor of Philosophy (Economics) Title: The origins and consequences of kin networks and marriage practices Examining Committee: Chair: Nicolas Schmitt Professor Gregory K. Dow Senior Supervisor Professor Alexander K. Karaivanov Supervisor Professor Erik O. Kimbrough Supervisor Associate Professor Argyros School of Business and Economics Chapman University Simon D. Woodcock Supervisor Associate Professor Chris Bidner Internal Examiner Associate Professor Siwan Anderson External Examiner Professor Vancouver School of Economics University of British Columbia Date Defended: July 31, 2018 ii Ethics Statement iii iii Abstract In the first chapter, I investigate a potential channel to explain the heterogeneity of kin networks across societies. I argue and test the hypothesis that female inheritance has historically had a posi- tive effect on in-marriage and a negative effect on female premarital relations and economic partic- ipation. In the second chapter, my co-authors and I provide evidence on the positive association of in-marriage and corruption. We also test the effect of family ties on nepotism in a bribery experi- ment. The third chapter presents my second joint paper on the consequences of kin networks. -

Performing Masculinity in Peri-Urban China: Duty, Family, Society
The London School of Economics and Political Science Performing Masculinity in Peri-Urban China: Duty, Family, Society Magdalena Wong A thesis submitted to the Department of Anthropology of the London School of Economics for the degree of Doctor of Philosophy, London December 2016 1 DECLARATION I certify that the thesis I have presented for examination for the MPhil/ PhD degree of the London School of Economics and Political Science is solely my own work other than where I have clearly indicated that it is the work of others (in which case the extent of any work carried out jointly by me and any other person is clearly identified in it). The copyright of this thesis rests with the author. Quotation from it is permitted, provided that full acknowledgement is made. This thesis may not be reproduced without my prior written consent. I warrant that this authorisation does not, to the best of my belief, infringe the rights of any third party. I declare that my thesis consists of 97,927 words. Statement of use of third party for editorial help I confirm that different sections of my thesis were copy edited by Tiffany Wong, Emma Holland and Eona Bell for conventions of language, spelling and grammar. 2 ABSTRACT This thesis examines how a hegemonic ideal that I refer to as the ‘able-responsible man' dominates the discourse and performance of masculinity in the city of Nanchong in Southwest China. This ideal, which is at the core of the modern folk theory of masculinity in Nanchong, centres on notions of men's ability (nengli) and responsibility (zeren). -

The Qadan, the Jebel Sahaba Cemetery and the Lithic
PL ISSN 0066–5924 INDEX 32597x (VAT 8%) DOI: 10.23858/APa58.2020 Institute of Archaeology and Ethnology Polish Academy of Sciences Archaeologia Polona vol. 58: 2020 Special theme: PREHistorY OF North-EAST AFRICA Volume dedicated to Prof. Michał Kobusiewicz on his 80th birthday Archaeologia Polona Volume 58 : 2020 Editor: DAGMARA H. WERRA Associate Editor MARZENA WOŹNY International Advisory Prof. PETER BOGUCKI – Princeton University, USA Board: Prof. MARTIN GOJDA – University of Western Bohemia, Pilsen, Czech Republic Prof. ANTHONY HARDING – University of Exeter, Great Britain Prof. MARIA IACOVOU – University of Cyprus, Nicosia, Cyprus Prof. CAROLA METZNER-NEBELSICK – University of Munich, Germany Prof. BJØRNAR OLSEN – University of Tromsø, Norway Prof. GUIDO VANNINI – University of Florence, Italy Editors of the volume: PRZEMYSłAW Bobrowski and MirosłAW MASOJć Linguistic consultation: PAUL BARFORD Cover illustration: MACIEJ JÓRDECZKA Reviewers: Mirosław Furmanek (Wroclaw), Elena Garcea (Cassino), Maria Gatto (Leicester), Bolesław Ginter (Cracow), Tomasz Herbich (Warsaw), Karla Kroeper (Berlin), Alice Leplongeon (Leuven), Maria Kaczmarek (Poznan), Andrea Manzo (Naples), Arkadiusz Marciniak (Poznan), Henryk Paner (Gdansk), Tomasz Płonka (Wroclaw), Włodzimierz Rączkowski (Poznan), Andrzej Rozwadowski (Poznan), Jiří Svoboda (Brno), Philip Van Peer (Leuven), András Zboray (Budapest). Editorial office: Institute of Archaeology and Ethnology Polish Academy of Sciences, 00-140 Warsaw; Al. Solidarności 105 Tel. (4822) 6202881 ext. 154; Fax. (4822) 624 01 00 E-mail: [email protected] Website: http://journals.iaepan.pl/apolona DOI: 10.23858/APa DOI: 10.23858/APa58.2020 Archaeologia Polona Copyright © 2020 by Institute of Archaeology and Ethnology Polish Academy of Sciences. All articles in this volume are published in an open access under the CC BY 4.0 license (https://creativecommons.org/licenses/by/4.0/). -

Chapter 4: Egypt, 3100 B.C
0066-0081 CH04-846240 10/24/02 5:57 PM Page 66 CHAPTER Egypt 4 3100 B.C.–671 B.C. Tutankhamen’s gold mask ᭤ ᭡ Wooden Egyptian sandals 2600 B.C. 2300 B.C. 1786 B.C. 1550 B.C. 671 B.C. Old Kingdom Middle Kingdom Hyksos invade Ahmose founds Assyrians take established begins Egypt the New over Egypt Kingdom 66 UNIT 2 RIVER VALLEY CIVILIZATIONS 0066-0081 CH04-846240 10/24/02 5:57 PM Page 67 Chapter Focus Read to Discover Chapter Overview Visit the Human Heritage Web site • Why the Nile River was so important to the growth of at humanheritage.glencoe.com Egyptian civilization. and click on Chapter 4—Chapter • How Egyptian religious beliefs influenced the Old Kingdom. Overviews to preview this chapter. • What happened during Egypt’s Middle Kingdom. • Why Egyptian civilization grew and then declined during the New Kingdom. • What the Egyptians contributed to other civilizations. Terms to Learn People to Know Places to Locate shadoof Narmer Nile River pharaoh Ahmose Punt pyramids Thutmose III Thebes embalming Hatshepsut mummy Amenhotep IV legend hieroglyphic papyrus Why It’s Important The Egyptians settled in the Nile River val- ley of northeast Africa. They most likely borrowed ideas such as writing from the Sumerians. However, the Egyptian civiliza- tion lasted far longer than the city-states and empires of Mesopotamia. While the people of Mesopotamia fought among themselves, Egypt grew into a rich, powerful, and unified kingdom. The Egyptians built a civilization that lasted for more than 2,000 years and left a lasting influence on the world. -

Daring to Care Reflections on Egypt Before the Revolution and the Way Forward
THE ASSOCIATION OF INTERNATIONAL CIVIL SERVANTS IN EGYPT Daring To Care Reflections on Egypt Before The Revolution And The Way Forward Experts’ Views On The Problems That Have Been Facing Egypt Throughout The First Decade Of The Millennium And Ways To Solve Them Daring to Care i Daring to Care ii Daring to Care Daring to Care Reflections on Egypt before the revolution and the way forward A Publication of the Association of International Civil Servants (AFICS-Egypt) Registered under No.1723/2003 with Ministry of Solidarity iii Daring to Care First published in Egypt in 2011 A Publication of the Association of International Civil Servants (AFICS-Egypt) ILO Cairo Head Office 29, Taha Hussein st. Zamalek, Cairo Registered under No.1723/2003 with Ministry of Solidarity Copyright © AFICS-Egypt All rights reserved Printed in Egypt All articles and essays appearing in this book as appeared in Beyond - Ma’baed publication in English or Arabic between 2002 and 2010. Beyond is the English edition, appeared quarterly as a supplement in Al Ahram Weekly newspaper. Ma’baed magazine is its Arabic edition and was published independently by AFICS-Egypt. BEYOND-MA’BAED is a property of AFICS EGYPT No part of this publication may be reproduced or transmitted or utilised in any form or by any means, electronic or mechanical, photocopying or otherwise, without prior permission of AFICS Egypt. Printed in Egypt by Moody Graphic International Ltd. 7, Delta st. ,Dokki 12311, Giza, Egypt - www.moodygraphic.com iv Daring to Care To those who have continuously worked at stirring the conscience of Egypt, reminding her of her higher calling and better self. -

Egyptian Literature
The Project Gutenberg EBook of Egyptian Literature This eBook is for the use of anyone anywhere at no cost and with almost no restrictions whatsoever. You may copy it, give it away or re-use it under the terms of the Project Gutenberg License included with this eBook or online at http://www.gutenberg.org/license Title: Egyptian Literature Release Date: March 8, 2009 [Ebook 28282] Language: English ***START OF THE PROJECT GUTENBERG EBOOK EGYPTIAN LITERATURE*** Egyptian Literature Comprising Egyptian Tales, Hymns, Litanies, Invocations, The Book Of The Dead, And Cuneiform Writings Edited And With A Special Introduction By Epiphanius Wilson, A.M. New York And London The Co-Operative Publication Society Copyright, 1901 The Colonial Press Contents Special Introduction. 2 The Book Of The Dead . 7 A Hymn To The Setting Sun . 7 Hymn And Litany To Osiris . 8 Litany . 9 Hymn To R ....................... 11 Hymn To The Setting Sun . 15 Hymn To The Setting Sun . 19 The Chapter Of The Chaplet Of Victory . 20 The Chapter Of The Victory Over Enemies. 22 The Chapter Of Giving A Mouth To The Overseer . 24 The Chapter Of Giving A Mouth To Osiris Ani . 24 Opening The Mouth Of Osiris . 25 The Chapter Of Bringing Charms To Osiris . 26 The Chapter Of Memory . 26 The Chapter Of Giving A Heart To Osiris . 27 The Chapter Of Preserving The Heart . 28 The Chapter Of Preserving The Heart . 29 The Chapter Of Preserving The Heart . 30 The Chapter Of Preserving The Heart . 30 The Heart Of Carnelian . 31 Preserving The Heart . 31 Preserving The Heart . -
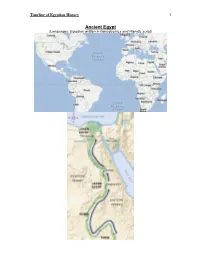
Timeline .Pdf
Timeline of Egyptian History 1 Ancient Egypt (Languages: Egyptian written in hieroglyphics and Hieratic script) Timeline of Egyptian History 2 Early Dynastic Period 3100–2686 BCE • 1st & 2nd Dynasty • Narmer aka Menes unites Upper & Lower Egypt • Hieroglyphic script developed Left: Narmer wearing the crown of Lower Egypt, the “Deshret”, or Red Crown Center: the Deshret in hieroglyphics; Right: The Red Crown of Lower Egypt Narmer wearing the crown of Upper Egypt, the “Hedjet”, or White Crown Center: the Hedjet in hieroglyphics; Right: The White Crown of Upper Egypt Pharaoh Djet was the first to wear the combined crown of Upper and Lower Egypt, the “Pschent” (pronounced Pskent). Timeline of Egyptian History 3 Old Kingdom 2686–2181 BCE • 3rd – 6th Dynasty • First “Step Pyramid” (mastaba) built at Saqqara for Pharaoh Djoser (aka Zoser) Left: King Djoser (Zoser), Righr: Step pyramid at Saqqara • Giza Pyramids (Khufu’s pyramid – largest for Pharaoh Khufu aka Cheops, Khafra’s pyramid, Menkaura’s pyramid – smallest) Giza necropolis from the ground and the air. Giza is in Lower Egypt, mn the outskirts of present-day Cairo (the modern capital of Egypt.) • The Great Sphinx built (body of a lion, head of a human) Timeline of Egyptian History 4 1st Intermediate Period 2181–2055 BCE • 7th – 11th Dynasty • Period of instability with various kings • Upper & Lower Egypt have different rulers Middle Kingdom 2055–1650 BCE • 12th – 14th Dynasty • Temple of Karnak commences contruction • Egyptians control Nubia 2nd Intermediate Period 1650–1550 BCE • 15th – 17th Dynasty • The Hyksos come from the Levant to occupy and rule Lower Egypt • Hyksos bring new technology such as the chariot to Egypt New Kingdom 1550–1069 BCE (Late Egyptian language) • 18th – 20th Dynasty • Pharaoh Ahmose overthrows the Hyksos, drives them out of Egypt, and reunites Upper & Lower Egypt • Pharaoh Hatshepsut, a female, declares herself pharaoh, increases trade routes, and builds many statues and monuments. -

Lukenya Hill, Kenya)
Late Pleistocene age and archaeological context for the hominin calvaria from GvJm-22 (Lukenya Hill, Kenya) Christian A. Tryona,1, Isabelle Crevecoeurb, J. Tyler Faithc, Ravid Ekshtaina, Joelle Nivensd, David Pattersone, Emma N. Mbuaf, and Fred Spoorg,h aDepartment of Anthropology, Harvard University, Cambridge, MA 02138; bUnité Mixte de Recherche 5199, de la Préhistoire à l’Actuel: Culture, Environnement, et Anthropologie, Centre National de la Recherche Scientifique, Université de Bordeaux, 33615 Talence, France; cArchaeology Program, School of Social Science, University of Queensland, Brisbane, QLD 4072, Australia; dDepartment of Anthropology, New York University, New York, NY 10003; eCenter for the Advanced Study of Hominid Paleobiology, Department of Anthropology, The George Washington University, Washington, DC 20052; fNational Museums of Kenya, Nairobi, Kenya 00100; gDepartment of Human Evolution, Max Planck Institute for Evolutionary Anthropology, D-04103, Leipzig, Germany; and hDepartment of Cell and Developmental Biology, University College London, WC1E 6BT London, United Kingdom Edited by Erik Trinkaus, Washington University, St. Louis, MO, and approved January 16, 2015 (received for review September 19, 2014) Kenya National Museums Lukenya Hill Hominid 1 (KNM-LH 1) is Hominid 1 (KNM-LH 1) partial calvaria from site GvJm-22 at a Homo sapiens partial calvaria from site GvJm-22 at Lukenya Hill, Lukenya Hill, Kenya, the only eastern African fossil hominin Kenya, associated with Later Stone Age (LSA) archaeological de- from a Last Glacial Maximum [LGM; 19–26.4 kya (19)] LSA posits. KNM-LH 1 is securely dated to the Late Pleistocene, and archaeological context. We construct a revised accelerator mass samples a time and region important for understanding the origins spectrometry (AMS) radiocarbon chronology built on 26 new of modern human diversity. -

Amarna Period Down to the Opening of Sety I's Reign
oi.uchicago.edu STUDIES IN ANCIENT ORIENTAL CIVILIZATION * NO.42 THE ORIENTAL INSTITUTE OF THE UNIVERSITY OF CHICAGO Thomas A. Holland * Editor with the assistance of Thomas G. Urban oi.uchicago.edu oi.uchicago.edu Internet publication of this work was made possible with the generous support of Misty and Lewis Gruber THE ROAD TO KADESH A HISTORICAL INTERPRETATION OF THE BATTLE RELIEFS OF KING SETY I AT KARNAK SECOND EDITION REVISED WILLIAM J. MURNANE THE ORIENTAL INSTITUTE OF THE UNIVERSITY OF CHICAGO STUDIES IN ANCIENT ORIENTAL CIVILIZATION . NO.42 CHICAGO * ILLINOIS oi.uchicago.edu Library of Congress Catalog Card Number: 90-63725 ISBN: 0-918986-67-2 ISSN: 0081-7554 The Oriental Institute, Chicago © 1985, 1990 by The University of Chicago. All rights reserved. Published 1990. Printed in the United States of America. oi.uchicago.edu TABLE OF CONTENTS List of M aps ................................ ................................. ................................. vi Preface to the Second Edition ................................................................................................. vii Preface to the First Edition ................................................................................................. ix List of Bibliographic Abbreviations ..................................... ....................... xi Chapter 1. Egypt's Relations with Hatti From the Amarna Period Down to the Opening of Sety I's Reign ...................................................................... ......................... 1 The Clash of Empires -

I) If\L /-,7\ .L Ii Lo N\ C, ' II Ii Abstract Approved: 1'
AN ABSTRACT OF THE THESIS OF Asaad AI-Saleh for the Master of Arts Degree In English presented on _------'I'--'I--'J:..=u:o...1VL.c2=0"--'0"-=S'------ _ Title: Mustafa Sadiq al-Rafii: A Non-recognized Voice in the Chorus ofthe Arabic Literary Revival i) If\l /-,7\ .L Ii lo n\ C, ' II Ii Abstract Approved: 1'. C". C ,\,,: 41-------<..<.LI-hY,-""lA""""","""I,--ft-'t _ '" I) Abstract Mustafa Sadiq al-Rafii, a modem Egyptian writer with classical style, is not studied by scholars of Arabic literature as are his contemporary liberals, such as Taha Hussein. This thesis provides a historical background and a brief literary survey that helps contextualize al-Rafii, the period, and the area he came from. AI-Rafii played an important role in the two literary and intellectual schools during the Arabic literary revival, which extended from the French expedition (1798-1801) to around the middle of the twentieth century. These two schools, known as the Old and the New, vied to shape the literature and thought of Egypt and other Arab countries. The former, led by al-Rafii, promoted a return to classical Arabic styles and tried to strengthen the Islamic identity of Egypt. The latter called for cutting off Egypt from its Arabic history and rejected the dominance and continuity of classical Arabic language. AI-Rafii contributed to the Revival by supporting a line ofthought that has not been favored by pro-Westernization governments, which made his legacy almost forgotten. Deriving his literature from the canon of Arabic language, culture, and history, al-Rafii produced a literature based on a revived version of classical Arabic literature, an accomplishment which makes him unique among modem Arab writers. -

Egypt and Nubia
9 Egypt and Nubia Robert Morkot THE, EGYPTIAN E,MPIRE,IN NUBIA IN THE LATE, BRONZE AGE (t.1550-l 070B CE) Introdu,ct'ion:sef-def,nit'ion an,d. the ,irnperi.ol con.cept in Egypt There can be litde doubt rhat the Egyptian pharaohs and the elite of the New I(ngdom viewed themselvesas rulers of an empire. This universal rule is clearly expressedin royal imagerv and terminology (Grimal f986). The pharaoh is styied asthe "Ruler of all that sun encircles" and from the mid-f 8th Dynasry the tides "I(ing of kings" and "Ruler of the rulers," with the variants "Lion" or "Sun of the Rulers," emphasizepharaoh's preeminence among other monarchs.The imagery of krngship is of the all-conquering heroic ruler subjecting a1lforeign lands.The lcingin human form smiteshis enemies.Or, asthe celestialconqueror in the form of the sphinx, he tramples them under foot. In the reigns of Amenhotep III and Akhenaten this imagery was exrended to the king's wife who became the conqueror of the female enemies of Egypt, appearing like her husbandin both human and sphinx forms (Morkot 1986). The appropriateter- minology also appeared; Queen Tiye became "Mistress of all women" and "Great of terror in the foreign lands." Empire, for the Egyptians,equals force - "all lands are under his feet." This metaphor is graphically expressedin the royal footstools and painted paths decorated \Mith images of bound foreign rulers, crushed by pharaoh as he walked or sar. This imagery and terminologv indicates that the Egyptian attirude to their empire was universally applied irrespective of the peoples or countries. -

Who Was Who at Amarna
1 Who was Who at Amarna Akhenaten’s predecessors Amenhotep III: Akhenaten’s father, who ruled for nearly 40 years during the peak of Egypt’s New Kingdom empire. One of ancient Egypt’s most prolific builders, he is also known for his interest in the solar cult and promotion of divine kingship. He was buried in WV22 at Thebes, his mummy later cached with other royal mummies in the Tomb of Amenhotep II (KV 35) in the Valley of the Kings. Tiye: Amenhotep III’s chief wife and the mother of Akhenaten. Her parents Yuya and Tjuyu were from the region of modern Akhmim in Egypt’s south. She may have lived out her later years at Akhetaten and died in the 14th year of Akhenaten’s reign. Funerary equipment found in the Amarna Royal Tomb suggests she was originally buried there, although her mummy was later moved to Luxor and is perhaps to be identified as the ‘elder lady’ from the KV35 cache. Akhenaten and his family Akhenaten: Son and successor of Amenhotep III, known for his belief in a single solar god, the Aten. He spent most of his reign at Akhetaten (modern Amarna), the sacred city he created for the Aten. Akhenaten died of causes now unknown in the 17th year of his reign and was buried in the Amarna Royal Tomb. His body was probably relocated to Thebes and may be the enigmatic mummy recovered in the early 20th century in tomb KV55 in the Valley of the Kings. Nefertiti: Akhenaten’s principal queen. Little is known of her background, although she may also have come from Akhmim.