Comparison of Oak and Sugar Maple Distribution and Regeneration in Central Illinois Upland Oak Forests
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Genetic and Phenotypic Characterization of Figured Wood in Poplar
Genetic and Phenotypic Characterization of Figured Wood in Poplar Youran Fan1,2, Keith Woeste1,2, Daniel Cassens1, Charles Michler1,2, Daniel Szymanski3, and Richard Meilan1,2 1Department of Forestry and Natural Resources, 2Hardwood Tree Improvement and Regeneration Center, and 3Department of Agronomy; Purdue University, West Lafayette, Indiana 47907 Abstract Materials and Methods When “Curly Aspen” (Populus canescens) was first Preliminary Results characterized in the early 1940’s[1], it attracted the attention from the wood-products industry because Genetically engineer commercially 1) Histological sections reveal that “Curly Aspen” has strong “Curly Aspen” produces an attractive veneer as a important trees to form figure. ray flecks (Fig. 10) but this is not likely to be responsible result of its figured wood. Birdseye, fiddleback and for the figure seen. quilt are other examples of figured wood that are 2) Of the 15 SSR primer pairs[6, 7, 8] tested, three have been commercially important[2]. These unusual grain shown to be polymorphic. Others are now being tested. patterns result from changes in cell orientation in Figure 6. Pollen collection. Branches of Figure 7. Pollination. Branches Ultimately, our genetic fingerprinting technique will allow “Curly Aspen” were “forced” to shed collected from a female P. alba us to distinguish “Curly Aspen” from other genotypes. the xylem. Although 50 years have passed since Figure 1. Birdseye in maple. pollen under controlled conditions. growing at Iowa State University’s finding “Curly Aspen”, there is still some question Rotary cut, three-piece book McNay Farm (south of Lucas, IA). 3) 17 jars of female P. alba branches have been pollinated match (origin: North America). -

Sugar Maple - Oak - Hickory Forest State Rank: S3 - Vulnerable
Sugar Maple - Oak - Hickory Forest State Rank: S3 - Vulnerable Mesic Forest (RMF): Sugar Maple - Oak - Hickory Forests are most occurrences of RMF diverse forests in central and eastern in Massachusetts are west Massachusetts where conditions, of the Connecticut River including nutrient richness, support Valley. The presence of Northern Hardwood species mixed with multiple species of species of Oak - Hickory Forests; hickories and oaks in SMOH is a main The herbaceous layer varies from sparse difference between these to intermittent, with sparse spring two types. Broad-leaved ephemerals that may include bloodroot or Woodland-sedge is close trout-lily. Later occurring species may to being an indicator of include wild geranium, herb Robert, wild SMOH. RMF is Rock outcrops in the spring in Sugar Maple - licorice, maidenhair fern, bottlebrush Oak - Hickory Forest area. Photo: Patricia characterized by very Swain, NHESP. grass, and white wood aster. Broad- dense herbaceous growth of spring leaved, semi-evergreen broad-leaved ephemerals; SMOH shares some of the Description: Sugar Maple - Oak - woodland-sedge is close to an indicator of species but with fewer individuals of Hickory Forests occur in or east of the the community. Witch hazel, hepaticas, fewer species. SMOH has evergreen Connecticut River Valley in and wild oats usually occur in transitions ferns, Christmas fern and wood ferns, that Massachusetts. They are associated with to surrounding forest types. RMF lack. Oak - Hickory Forests and outcrops of circumneutral rock and slopes Dry, Rich Oak Forests/Woodlands lack below them that have more nutrients than abundant sugar maple, basswood, and are available in the surrounding forest. -
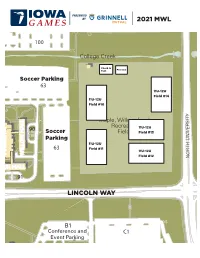
2021 Mwl 2 L
Schilletter - University SCHILLETTER - UNIVERSITY VILLAGE201R COMMUNITY 201B 201D 201N CENTER D A O APPLIED R GOLF COURSE SCIENCE 201P S MAINTENANCE T COMPLEX IV T OFFICE O T LONG ROAD S G DR NBUR IVE 201M BLANK201CE 201K 104 CLUBHOUSE APPLIED D SCIENCE A COMPLEX I O R 201L 102 G 201C APPLIED 103 N O SCIENCE L COMPLEX III APPLIED D SCIENCE 201J A O COMPLEX II 201H R WASTE S T CHEMICAL T 201F D 201G O HANDLING T 101 A S SENSITIVE BUILDING Veenker Memorial Golf Course O INSTRUMENT R FACILITY E LYNN G FUHRER N A LODGE T S 205 201E 201D 201B ISU FAMILY 200 RESOURCE CENTER 205 RIVE 201A BRUNER D Iow ay C reek D A O R L L O H Io C w S ay Cre ek Disc Golf Course 112 L 13 Veenker Memorial Golf Course TH S TRE Furnam Aquatic Center ET EXTENSION 4-H (City Of Ames) T 125 REE YOUTH BUILDING H ST 13T 112 J 124A 112 A 112 J 112 H EH&S ONTARIO STREET 122A ONTARIO STREET SERVICES 120A ROAD STANGE BUILDING 124 FRED115ERIKSEN 112 K 121 BLDG SERVICES COURT ADMINISTRATIVE 112 G HAWTHORN COURT COMMUNITY FREDERIKSEN WANDA DALEY DRIVE DRIVE CENTER COURT 112 K 112 K 122 LIBRARY 120 DOE STORAGE WAREHOUSE FACILITY 119 DOE DOE CONST D 112 D-1 MECH A DOE O R 29 MAINT DOE R PRINTING & E SHOP B ROY J. CARVER PUBLICATIONS31 A CO-LABORATORY BUILDING H 112N 29 NORTH CHILLED 112 B 112 F WATER PLANT ear Creek 112 C 112 D-2 Cl 28 28 Union Pacific Railroad 35 30 27 28A 33 Pammel Woods 12 MOLECULAR 28A FIRE SERVICE BIOLOGY BUILDING COMMUNICATIONS 32 A BUILDING METALS TRANSPORTATION S ADVANCED BUILDING 28A 110 DEVELOPMENT C RUMINANT I 32 TEACHING & HORSE BARN BUILDING T NUTRITION SERVICES E RESEARCH 79 STANGE ROAD STANGE N LAB Cemetery E BUILDING WINLOCK ROAD G LABORATORY MACH ADVANCED LAB SYSTEMS 11W 11E PAMMEL DRIVE NORTH UNIVERSITY BOULEVARD PAMMEL DRIVE SPEDDING WILHELM NATIONAL NATIONAL HYLAND AVENUE HYLAND 11 22 HACH HALL HALL HALL SCIENCE SWINE RSRCH LAB FOR AG. -

Folivory of Vine Maple in an Old-Growth Douglas-Fir-Western Hemlock Forest
3589 David M. Braun, Bi Runcheng, David C. Shaw, and Mark VanScoy, University of Washington, Wind River Canopy Crane Research Facility, 1262 Hemlock Rd., Carson, Washington 98610 Folivory of Vine Maple in an Old-growth Douglas-fir-Western Hemlock Forest Abstract Folivory of vine maple was documented in an old-growth Douglas-fir-western hemlock forest in southwest Washington. Leaf consumption by lepidopteran larvae was estimated with a sample of 450 tagged leaves visited weekly from 7 May to 11 October, the period from bud break to leaf drop. Lepidopteran taxa were identified by handpicking larvae from additional shrubs and rearing to adult. Weekly folivory peaked in May at 1.2%, after which it was 0.2% to 0.7% through mid October. Cumulative seasonal herbivory was 9.9% of leaf area. The lepidopteran folivore guild consisted of at least 22 taxa. Nearly all individuals were represented by eight taxa in the Geometridae, Tortricidae, and Gelechiidae. Few herbivores from other insect orders were ob- served, suggesting that the folivore guild of vine maple is dominated by these polyphagous lepidopterans. Vine maple folivory was a significant component of stand folivory, comparable to — 66% of the folivory of the three main overstory conifers. Because vine maple is a regionally widespread, often dominant understory shrub, it may be a significant influence on forest lepidopteran communities and leaf-based food webs. Introduction tract to defoliator outbreaks, less is known about endemic populations of defoliators and low-level Herbivory in forested ecosystems consists of the folivory. consumption of foliage, phloem, sap, and live woody tissue by animals. -

4-H Wood Science Leader Guide Glossary of Woodworking Terms
4-H Wood Science Leader Guide Glossary of Woodworking Terms A. General Terms B. Terms Used in the Lumber Industry d—the abbreviation for “penny” in designating nail boards—Lumber less than 2 inches in nominal size; for example, 8d nails are 8 penny nails, 2½” long. thickness and 1 inch and wider in width. fiber—A general term used for any long, narrow cell of board foot—A measurement of wood. A piece of wood wood or bark, other than vessels. that is 1 foot long by 1 foot wide by 1 inch thick. It can also be other sizes that have the same total amount grain direction—The direction of the annual rings of wood. For example, a piece of wood 2 feet long, showing on the face and sides of a piece of lumber. 6 inches wide, and 1 inch thick; or a piece 1 foot long, hardwood—Wood from a broad leaved tree and 6 inches wide, and 2 inches thick would also be 1 board characterized by the presence of vessels. (Examples: foot. To get the number of board feet in a piece of oak, maple, ash, and birch.) lumber, measure your lumber and multiply Length (in feet) x Width (in feet) x Thickness (in inches). The heartwood—The older, harder, nonliving portion of formula is written: wood. It is usually darker, less permeable, and more durable than sapwood. T” x W’ x L’ T” x W’ x L’ = Board feet or = Board feet 12 kiln dried—Wood seasoned in a humidity and temperature controlled oven to minimize shrinkage T” x W’ x L’ or = Board feet and warping. -

Self-Guided Neighborhood Tree Tour Created by Bhopesh Bassi
BELLEVUE NEIGHBORHOOD TREE AMBASSADOR PROGRAM Self-Guided Neighborhood Tree Tour Created by Bhopesh Bassi Neighborhood: Kelsey Creek Park and Farm Starting point: Kelsey Creek Park and Farm parking lot at 410 130th Place SE Summary/Theme: Introduction to plant and animal ecosystem in and around Kelsey Creek Park and Farm. Kelsey Creek Farm Park in Bellevue is a largely overlooked gem. While the park is tucked in the heart of neighborhoods, it feels like an isolated country woods experience. You may have the impression of having traveled back in time as you move from the modern parking lot to a more rural feel as you hike into the forest. Kelsey Creek is Seattle's largest watershed. It runs right down the middle of the park, providing habitat for spawning Salmon, Peamouth and for an abundant population of migratory and resident birds. All lifeform here depends on and contributes to each other and the park is a rich ecosystem. Bring your binoculars along if you also want to watch birds on this tour. BELLEVUE NEIGHBORHOOD TREE AMBASSADOR PROGRAM This tree tour was developed by one of Bellevue’s Neighborhood Tree Ambassador volunteers. The goal of the Neighborhood Tree Ambassador program is to help build community support for trees in Bellevue. Trees are an important part of our community because they provide significant health and environmental benefits. Trees: • Remove pollutants from the air and water • Reduce stress and improve focus • Lower air temperature • Pull greenhouse gases from the atmosphere • Reduce flooding and erosion caused by rain Bellevue has a goal to achieve a 40% tree canopy across the entire city. -

Veneer Guidelines 1
V T INDUSTRIES VENEER GUIDELINES 1 VTWO O D VENEERS PLAN I SLICED NATURAL MAPLE N ATURAL VARIATIONS The word natural brings to mind certain connotations like “beauty”, “warmth” and “purity”. Merriam-Webster defines natural as “occurring in conformity with the ordinary course of nature (the genetically controlled qualities of an organism): not marvelous or supernatural”. Wood is a product of nature, and in some cases, will accentuate and enhance a project design when used in its purest, or natural, state. However, as a product of nature, each wood species has certain intrinsic and industry-acceptable characteristics, which can vary from tree to tree and flitch (half log) to flitch. It is precisely these naturally occurring variations that provide such richness and uniqueness to each project design. Certain wood species such as natural maple and birch can vary widely in color range, which is why in many cases select white is specified so that the sapwood can be accumulated and spliced together to create a consistent color. The photos and information in this brochure are designed to assist you in specifying and receiving the product you envision. When specifying "natural" maple and birch, the veneer will contain unlimited amounts of Sapwood (the light portion of the log) and/or Heartwood (the dark portion of the log) unselected for color. If a light colored veneer is preferred, specify Select White (all Sapwood) maple or birch. If a dark colored veneer is preferred, specify Select Red/Brown (all Heartwood) Note, availability may be limited. 2 VTWO O D VENEERS PLAN I SLICED SELECT WHITE MAPLE HO W TO SPECIFY Natural veneers, such as maple and birch, may contain sapwood/ heartwood combinations, color streaks, spots and color variation from almost white to very dark. -

Street Tree Planting Procedures
Seattle Department of Transportation CLIENT ASSISTANCE MEMO 700 Fifth Avenue, Suite 2300 | P.O. Box 34996 Seattle, Washington 98124-4996 (206) 684-5253 | [email protected] 2300 SEATTLE PERMITS - Part of a multi- departmental City of Seattle series on getting a permit STREET TREE PLANTING PROCEDURES www.seattle.gov/transportation Last Revised 12/11/17 • Plant all small scale trees between 20 and 25 feet apart as measured from the center of the The City of Seattle encourages the planting of trees tree. Plant all upright or columnar trees between along public streets. A property owner may plant 15 and 25 feet apart. Plant all medium scale a tree in a planting strip if he or she first obtains a trees between 25 and 35 feet apart. permit from the City Arborist’s Office. • Trees must be planted to the following standards: There is no charge for the permit, but the City - 3½ feet back from the face of the curb. Arborist must approve the type of tree and the - 5 feet from underground utility lines. planting location. After you receive the permit to - 10 feet from power poles (15 feet plant, you will be responsible for properly planting recommended) and maintaining the tree in perpetuity. This includes - 7½ feet from driveways (10 feet watering during the drier seasons, mulching and recommended) pruning. - 20 feet from street lights or other existing trees. Besides selecting a tree for aesthetics, please - 30 feet from street intersections. consider the following before you plant: • In most cases, the Arborist only approves the PROHIBITED STREET TREES FOR SEATTLE planting of trees when there is a curbed roadway City Ordinance No. -

Oak Willow Cherry, Plum Blueberry, Cranberry
Top 10 OAK Family: Fagaceae Genus: Quercus The mighty oak supports 534 species of butterflies/moths, and its acorns feed deer, 1 turkeys, bear, squirrels, even wood ducks. There are 80 oak species in North America. WILLOW BLUEBERRY, Family: Salicaceae CRANBERRY 2 Genus: Salix Family: Ericaceae 7 Genus: Vaccinium With 97 willow species in north america, there are lots to choose Underused in ornamental plant- from. they support some of the ings, they host butterflies/moths showiest butterflies, e.g., viceroys. and nourish birds and mammals. CHERRY, PLUM MAPLE Family: Rosaceae Family: Aceraceae 3 Genus: Prunus Genus: Acer 8 native plums and wild cherries, since the demise of the american including black cherry, choke- chestnut, maples have expanded cherry, and pin cherry, are excel- their role in the forest. they sup- lent sources of food for wildlife. port 285 lepidoptera species. BIRCH ELM Family: Betulaceae Family: Ulmaceae 4 Genus: Betula Genus: Ulmus 9 birches support hundreds of intense breeding programs have butterfly/moth species, plus produced disease-resistant ameri- they produce seeds and flower can elms. a few caterpillars eat buds that feed a variety of birds, nothing else and look like its leaves. from songbirds to turkeys. POPLAR PINE Family: Salicaceae Family: Pinaceae 5 Genus: Populus Genus: Pinus 10 lepidoptera love these fast-growing despite their resins and terpenes, trees. they harbor 7 giant silk moth, pine needles are a favorite food 7 sphinx moth, 77 noctuid (noctur- of 203 species of butterflies/ nal moth) and 10 butterfly species. moths in eastern forests. CRABAPPLE This hornworm is the larva of the hummingbird sphinx Family: Rosaceae moth. -

Woodworking Glossary, a Comprehensive List of Woodworking Terms and Their Definitions That Will Help You Understand More About Woodworking
Welcome to the Woodworking Glossary, a comprehensive list of woodworking terms and their definitions that will help you understand more about woodworking. Each word has a complete definition, and several have links to other pages that further explain the term. Enjoy. Woodworking Glossary A | B | C | D | E | F | G | H | I | J | K | L | M | N | O | P | Q | R | S | T | U | V | W | X | Y | Z | #'s | A | A-Frame This is a common and strong building and construction shape where you place two side pieces in the orientation of the legs of a letter "A" shape, and then cross brace the middle. This is useful on project ends, and bases where strength is needed. Abrasive Abrasive is a term use to describe sandpaper typically. This is a material that grinds or abrades material, most commonly wood, to change the surface texture. Using Abrasive papers means using sandpaper in most cases, and you can use it on wood, or on a finish in between coats or for leveling. Absolute Humidity The absolute humidity of the air is a measurement of the amount of water that is in the air. This is without regard to the temperature, and is a measure of how much water vapor is being held in the surrounding air. Acetone Acetone is a solvent that you can use to clean parts, or remove grease. Acetone is useful for removing and cutting grease on a wooden bench top that has become contaminated with oil. Across the Grain When looking at the grain of a piece of wood, if you were to scratch the piece perpendicular to the direction of the grain, this would be an across the grain scratch. -

A Glossary of Common Forestry Terms
W 428 A Glossary of Common Forestry Terms A Glossary of Common Forestry Terms David Mercker, Extension Forester University of Tennessee acre artificial regeneration A land area of 43,560 square feet. An acre can take any shape. If square in shape, it would measure Revegetating an area by planting seedlings or approximately 209 feet per side. broadcasting seeds rather than allowing for natural regeneration. advance reproduction aspect Young trees that are already established in the understory before a timber harvest. The compass direction that a forest slope faces. afforestation bareroot seedlings Establishing a new forest onto land that was formerly Small seedlings that are nursery grown and then lifted not forested; for instance, converting row crop land without having the soil attached. into a forest plantation. AGE CLASS (Cohort) The intervals into which the range of tree ages are grouped, originating from a natural event or human- induced activity. even-aged A stand in which little difference in age class exists among the majority of the trees, normally no more than 20 percent of the final rotation age. uneven-aged A stand with significant differences in tree age classes, usually three or more, and can be basal area (BA) either uniformly mixed or mixed in small groups. A measurement used to help estimate forest stocking. Basal area is the cross-sectional surface area (in two-aged square feet) of a standing tree’s bole measured at breast height (4.5 feet above ground). The basal area A stand having two distinct age classes, each of a tree 14 inches in diameter at breast height (DBH) having originated from separate events is approximately 1 square foot, while an 8-inch DBH or disturbances. -

CFD Based Analysis of Swietenia Mahogani Seed and Its Applications
IAETSD JOURNAL FOR ADVANCED RESEARCH IN APPLIED SCIENCES ISSN NO: 2394-8442 CFD Based Analysis of Swietenia Mahogani Seed and its Applications Titu Wilson*1, Anandu Raj *2, Susan K Thomas#3 * Department of Aeronautical Engineering, Mount Zion College of Engineering, Pathanamthitta, Kerala, INDIA, # Assistant Professor, Department of Aeronautical Engineering, Mount Zion College of Engineering, Pathanamthitta, Kerala, INDIA [email protected] [email protected] [email protected] Abstract— Swietenia Mahogani (Mahogany) seeds shows an extraordinary falling characteristic known as autorotation. The mahogany seeds are equipped with a small wing, which starts to auto rotate within 1 meter after they are detached from the tree. The seeds rotates in its axis centre similar to that of the gyration of propeller about its pivot point. This motion generates lift which slows down the falling speed and provides an additional time to stay afloat. In this paper the flow field around a mahogany seed under free fall condition is analyzed to discover its aerodynamic characteristics and study about its potential use as a wind turbine. Keywords— Autorotation, Axis centre, Gyration, Pivot point, Swietenia Mahagoni, Wind turbine. I. INTRODUCTION Nature is the best fascinating source of creativity and inspiration. Nature can inspire us about systems structures, designs, materials, processes, aesthetics, etc. Biomimetics is the term used to describe the substances, equipment, mechanism, and systems by humans to imitate natural systems and designs, aiming at solving problems by first examining, and then imitating or drawing inspiration from models in nature [1,2,3]. Mahogany seeds are one of the most generally interesting specimens for biological aerodynamic study within nature.