Characterization of Vitreous and Aqueous Proteome in Humans With
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Insulin-Like Growth Factor Binding Protein-3 Exerts Its Anti-Metastatic Effect in Aerodigestive Tract Cancers by Disrupting the Protein Stability of Vimentin
cancers Article Insulin-Like Growth Factor Binding Protein-3 Exerts Its Anti-Metastatic Effect in Aerodigestive Tract Cancers by Disrupting the Protein Stability of Vimentin Huong Thuy Le 1,†,‡, Ho Jin Lee 1,†, Jaebeom Cho 1, Hye-Young Min 1, Ji-Sun Lee 1, Su-Jae Lee 2 and Ho-Young Lee 1,* 1 Creative Research Initiative Center for Concurrent Control of Emphysema and Lung Cancer, College of Pharmacy and Research Institute of Pharmaceutical Sciences, Seoul National University, Seoul 08826, Korea; [email protected] (H.T.L.); [email protected] (H.J.L.); [email protected] (J.C.); [email protected] (H.-Y.M.); [email protected] (J.-S.L.) 2 Department of Life Science, Research Institute for Natural Sciences, Hanyang University, Seoul 04763, Korea; [email protected] * Correspondence: [email protected]; Tel.: +82-2-880-9277; Fax: +82-2-6280-5327 † These two authors contributed equally to this work and should be considered as first author. ‡ Current Address: Faculty of Pharmacy, Ton Duc Thang University, Ho Chi Minh City 700000, Vietnam. Simple Summary: Local invasion and distal metastasis are the main causes of cancer-related death and the poor prognosis of patients with aerodigestive tract cancers. Therefore, understanding the biology of invasion and metastasis is important for the development of effective therapeutic strategies. The present study shows that insulin-like growth factor binding protein-3 (IGFBP-3) inhibits the migration and invasion of non-small cell lung cancer (NSCLC) and head and neck squamous cell carcinoma (HNSCC) cells in vitro and the development of metastasized tumors in vivo. -

With Focus on the Genus Handleyomys and Related Taxa
Brigham Young University BYU ScholarsArchive Theses and Dissertations 2015-04-01 Evolution and Biogeography of Mesoamerican Small Mammals: With Focus on the Genus Handleyomys and Related Taxa Ana Villalba Almendra Brigham Young University - Provo Follow this and additional works at: https://scholarsarchive.byu.edu/etd Part of the Biology Commons BYU ScholarsArchive Citation Villalba Almendra, Ana, "Evolution and Biogeography of Mesoamerican Small Mammals: With Focus on the Genus Handleyomys and Related Taxa" (2015). Theses and Dissertations. 5812. https://scholarsarchive.byu.edu/etd/5812 This Dissertation is brought to you for free and open access by BYU ScholarsArchive. It has been accepted for inclusion in Theses and Dissertations by an authorized administrator of BYU ScholarsArchive. For more information, please contact [email protected], [email protected]. Evolution and Biogeography of Mesoamerican Small Mammals: Focus on the Genus Handleyomys and Related Taxa Ana Laura Villalba Almendra A dissertation submitted to the faculty of Brigham Young University in partial fulfillment of the requirements for the degree of Doctor of Philosophy Duke S. Rogers, Chair Byron J. Adams Jerald B. Johnson Leigh A. Johnson Eric A. Rickart Department of Biology Brigham Young University March 2015 Copyright © 2015 Ana Laura Villalba Almendra All Rights Reserved ABSTRACT Evolution and Biogeography of Mesoamerican Small Mammals: Focus on the Genus Handleyomys and Related Taxa Ana Laura Villalba Almendra Department of Biology, BYU Doctor of Philosophy Mesoamerica is considered a biodiversity hot spot with levels of endemism and species diversity likely underestimated. For mammals, the patterns of diversification of Mesoamerican taxa still are controversial. Reasons for this include the region’s complex geologic history, and the relatively recent timing of such geological events. -

Evolutionary History of Human Plasmodium Vivax Revealed by Genome-Wide Analyses of Related Ape Parasites
Evolutionary history of human Plasmodium vivax revealed by genome-wide analyses of related ape parasites Dorothy E. Loya,b,1, Lindsey J. Plenderleithc,d,1, Sesh A. Sundararamana,b, Weimin Liua, Jakub Gruszczyke, Yi-Jun Chend,f, Stephanie Trimbolia, Gerald H. Learna, Oscar A. MacLeanc,d, Alex L. K. Morganc,d, Yingying Lia, Alexa N. Avittoa, Jasmin Gilesa, Sébastien Calvignac-Spencerg, Andreas Sachseg, Fabian H. Leendertzg, Sheri Speedeh, Ahidjo Ayoubai, Martine Peetersi, Julian C. Raynerj, Wai-Hong Thame,f, Paul M. Sharpc,d,2, and Beatrice H. Hahna,b,2,3 aDepartment of Medicine, University of Pennsylvania, Philadelphia, PA 19104; bDepartment of Microbiology, University of Pennsylvania, Philadelphia, PA 19104; cInstitute of Evolutionary Biology, University of Edinburgh, Edinburgh EH9 3FL, United Kingdom; dCentre for Immunity, Infection and Evolution, University of Edinburgh, Edinburgh EH9 3FL, United Kingdom; eWalter and Eliza Hall Institute of Medical Research, Parkville VIC 3052, Australia; fDepartment of Medical Biology, The University of Melbourne, Parkville VIC 3010, Australia; gRobert Koch Institute, 13353 Berlin, Germany; hSanaga-Yong Chimpanzee Rescue Center, International Development Association-Africa, Portland, OR 97208; iRecherche Translationnelle Appliquée au VIH et aux Maladies Infectieuses, Institut de Recherche pour le Développement, University of Montpellier, INSERM, 34090 Montpellier, France; and jMalaria Programme, Wellcome Trust Sanger Institute, Genome Campus, Hinxton Cambridgeshire CB10 1SA, United Kingdom Contributed by Beatrice H. Hahn, July 13, 2018 (sent for review June 12, 2018; reviewed by David Serre and L. David Sibley) Wild-living African apes are endemically infected with parasites most recently in bonobos (Pan paniscus)(7–11). Phylogenetic that are closely related to human Plasmodium vivax,aleadingcause analyses of available sequences revealed that ape and human of malaria outside Africa. -

Vertebrate Fatty Acid and Retinoid Binding Protein Genes and Proteins: Evidence for Ancient and Recent Gene Duplication Events
In: Advances in Genetics Research. Volume 11 ISBN: 978-1-62948-744-1 Editor: Kevin V. Urbano © 2014 Nova Science Publishers, Inc. Chapter 7 Vertebrate Fatty Acid and Retinoid Binding Protein Genes and Proteins: Evidence for Ancient and Recent Gene Duplication Events Roger S. Holmes Eskitis Institute for Drug Discovery and School of Biomolecular and Physical Sciences, Griffith University, Nathan, QLD, Australia Abstract Fatty acid binding proteins (FABP) and retinoid binding proteins (RBP) are members of a family of small, highly conserved cytoplasmic proteins that function in binding and facilitating the cellular uptake of fatty acids, retinoids and other hydrophobic compounds. Several human FABP-like genes are expressed in the body: FABP1 (liver); FABP2 (intestine); FABP3 (heart and skeletal muscle); FABP4 (adipocyte); FABP5 (epidermis); FABP6 (ileum); FABP7 (brain); FABP8 (nervous system); FABP9 (testis); and FABP12 (retina and testis). A related gene (FABP10) is expressed in lower vertebrate liver and other tissues. Four RBP genes are expressed in human tissues: RBP1 (many tissues); RBP2 (small intestine epithelium); RBP5 (kidney and liver); and RBP7 (kidney and heart). Comparative FABP and RBP amino acid sequences and structures and gene locations were examined using data from several vertebrate genome projects. Sequence alignments, key amino acid residues and conserved predicted secondary and tertiary structures were also studied, including lipid binding regions. Vertebrate FABP- and RBP- like genes usually contained 4 coding exons in conserved locations, supporting a common evolutionary origin for these genes. Phylogenetic analyses examined the relationships and evolutionary origins of these genes, suggesting division into three FABP gene classes: 1: FABP1, FABP6 and FABP10; 2: FABP2; and 3, with 2 groups: 3A: FABP4, FABP8, FABP9 and FABP12; and 3B: and FABP3, FABP5 and FABP7. -

Supplementary Table 2
Supplementary Table 2. Differentially Expressed Genes following Sham treatment relative to Untreated Controls Fold Change Accession Name Symbol 3 h 12 h NM_013121 CD28 antigen Cd28 12.82 BG665360 FMS-like tyrosine kinase 1 Flt1 9.63 NM_012701 Adrenergic receptor, beta 1 Adrb1 8.24 0.46 U20796 Nuclear receptor subfamily 1, group D, member 2 Nr1d2 7.22 NM_017116 Calpain 2 Capn2 6.41 BE097282 Guanine nucleotide binding protein, alpha 12 Gna12 6.21 NM_053328 Basic helix-loop-helix domain containing, class B2 Bhlhb2 5.79 NM_053831 Guanylate cyclase 2f Gucy2f 5.71 AW251703 Tumor necrosis factor receptor superfamily, member 12a Tnfrsf12a 5.57 NM_021691 Twist homolog 2 (Drosophila) Twist2 5.42 NM_133550 Fc receptor, IgE, low affinity II, alpha polypeptide Fcer2a 4.93 NM_031120 Signal sequence receptor, gamma Ssr3 4.84 NM_053544 Secreted frizzled-related protein 4 Sfrp4 4.73 NM_053910 Pleckstrin homology, Sec7 and coiled/coil domains 1 Pscd1 4.69 BE113233 Suppressor of cytokine signaling 2 Socs2 4.68 NM_053949 Potassium voltage-gated channel, subfamily H (eag- Kcnh2 4.60 related), member 2 NM_017305 Glutamate cysteine ligase, modifier subunit Gclm 4.59 NM_017309 Protein phospatase 3, regulatory subunit B, alpha Ppp3r1 4.54 isoform,type 1 NM_012765 5-hydroxytryptamine (serotonin) receptor 2C Htr2c 4.46 NM_017218 V-erb-b2 erythroblastic leukemia viral oncogene homolog Erbb3 4.42 3 (avian) AW918369 Zinc finger protein 191 Zfp191 4.38 NM_031034 Guanine nucleotide binding protein, alpha 12 Gna12 4.38 NM_017020 Interleukin 6 receptor Il6r 4.37 AJ002942 -

Supplementary Table 1
Supplementary Table 1. 492 genes are unique to 0 h post-heat timepoint. The name, p-value, fold change, location and family of each gene are indicated. Genes were filtered for an absolute value log2 ration 1.5 and a significance value of p ≤ 0.05. Symbol p-value Log Gene Name Location Family Ratio ABCA13 1.87E-02 3.292 ATP-binding cassette, sub-family unknown transporter A (ABC1), member 13 ABCB1 1.93E-02 −1.819 ATP-binding cassette, sub-family Plasma transporter B (MDR/TAP), member 1 Membrane ABCC3 2.83E-02 2.016 ATP-binding cassette, sub-family Plasma transporter C (CFTR/MRP), member 3 Membrane ABHD6 7.79E-03 −2.717 abhydrolase domain containing 6 Cytoplasm enzyme ACAT1 4.10E-02 3.009 acetyl-CoA acetyltransferase 1 Cytoplasm enzyme ACBD4 2.66E-03 1.722 acyl-CoA binding domain unknown other containing 4 ACSL5 1.86E-02 −2.876 acyl-CoA synthetase long-chain Cytoplasm enzyme family member 5 ADAM23 3.33E-02 −3.008 ADAM metallopeptidase domain Plasma peptidase 23 Membrane ADAM29 5.58E-03 3.463 ADAM metallopeptidase domain Plasma peptidase 29 Membrane ADAMTS17 2.67E-04 3.051 ADAM metallopeptidase with Extracellular other thrombospondin type 1 motif, 17 Space ADCYAP1R1 1.20E-02 1.848 adenylate cyclase activating Plasma G-protein polypeptide 1 (pituitary) receptor Membrane coupled type I receptor ADH6 (includes 4.02E-02 −1.845 alcohol dehydrogenase 6 (class Cytoplasm enzyme EG:130) V) AHSA2 1.54E-04 −1.6 AHA1, activator of heat shock unknown other 90kDa protein ATPase homolog 2 (yeast) AK5 3.32E-02 1.658 adenylate kinase 5 Cytoplasm kinase AK7 -

1 Evolution of Sexual Development and Sexual Dimorphism in Insects 1
1 Evolution of sexual development and sexual dimorphism in insects 2 3 Ben R. Hopkins1* & Artyom Kopp1 4 5 1. Department of Evolution and Ecology, University of California – Davis, Davis, USA 6 7 * Corresponding author 8 9 Corresponding author email: [email protected] 10 11 12 13 14 Abstract 15 Most animal species consist of two distinct sexes. At the morphological, physiological, and 16 behavioural levels the differences between males and females are numerous and dramatic, yet 17 at the genomic level they are often slight or absent. This disconnect is overcome because simple 18 genetic differences or environmental signals are able to direct the sex-specific expression of a 19 shared genome. A canonical picture of how this process works in insects emerged from decades 20 of work on Drosophila. But recent years have seen an explosion of molecular-genetic and 21 developmental work on a broad range of insects. Drawing these studies together, we describe 22 the evolution of sexual dimorphism from a comparative perspective and argue that insect sex 23 determination and differentiation systems are composites of rapidly evolving and highly 24 conserved elements. 25 1 26 Introduction 27 Anisogamy is the definitive sex difference. The bimodality in gamete size it describes 28 represents the starting point of a cascade of evolutionary pressures that have generated 29 remarkable divergence in the morphology, physiology, and behaviour of the sexes [1]. But 30 sexual dimorphism presents a paradox: how can a genome largely shared between the sexes 31 give rise to such different forms? A powerful resolution is via sex-specific expression of shared 32 genes. -

Dynamic Approaches to Improve Glycemic Control and Primary Diabetes Care 1‑Or 2‑Or 3‑Or
DYNAMIC APPROACHES TO IMPROVE GLYCEMIC CONTROL AND PRIMARY DIABETES CARE DYNAMIC APPROACHES TO IMPROVE GLYCEMIC Figure. CONTROL AND PRIMARY DIABETES CARE 1‑OR Characterizing Clinical Inertia in a Large, National Database CORI R. RATTELMAN, ANUPAMA ARORA, JOHN K. CUDDEBACK, ELIZABETH L. CIEMINS, Alexandria, VA Objective: To characterize clinical inertia in the treatment of diabetes using a large, geographically diverse clinical database. Study Design: A retrospective descriptive analysis was conducted in a clinical database containing 22 million patient records across 22 health care organizations (HCOs). Population Studied: A total of 281,000 patients aged 18-75 were included during the 5.5-year study period (1/2012-6/2017). Patients had an outpatient visit in the last 12 months of the study period, an HbA1c in the last 24-30 months (index A1c), and a diagnosis of type 2 DM on a claim or EHR prob- lem list at least 6 months prior to index A1c. A subset of 47,693 patients with an index A1c ≥8 and a prior A1c ≥8 or lack thereof, was observed for Supported By: National Institute of Diabetes and Digestive and Kidney Diseases four 6-month follow-up periods for actions including a new class of diabetes (T35DK104689); Yale University medication prescribed or an A1c <8. The absence of observable action fol- lowing index A1c suggests potential “clinical inertia.” Principal Findings: Six months following an index A1c≥8, 55% of patients 3‑OR received no observable clinical action ranging from 45-65% across HCOs Influence of Diabetes Complications on the Cost‑Effectiveness of and 18-96% across individual providers. -

Inhibition of the DNA-Binding Activity of Drosophila Suppressor of Hairless and of Its Human Homolog, KBF2/RBP-JK, by Direct
Downloaded from genesdev.cshlp.org on October 1, 2021 - Published by Cold Spring Harbor Laboratory Press Inhibition of the DNA-binding activity of Drosophila Suppressor of Hairless and of its human homolog, KBF2/RBP-JK, by direct protein-protein interaction with Drosophila Hairless Christel Brou/'^ Frederique Logeat,^'^ Magalie Lecourtois,^ Joel Vandekerckhove,^ Philippe Kourilsky/ Francois Schweisguth/ and Alain Israel^'^ ^Unite de Biologic Moleculaire de I'Expression Genique, URA 1149 Centre National de la Recherche Scientifique (CNRS), Institut Pasteur, 75724 Paris cedex 15, France; ^Institut Jacques Monod, CNRS Universite Paris VII, 75251 Paris cedex 05, France; ^Laboratorium voor Genetica, Rijksuniversiteit Gent, B-9000 Gent, Belgium; '^Unite de Biologic Moleculaire du Gene, U. 277 Institut National de la Sante et de la Recherche Medicale (INSERM), Institut Pasteur, 75724 Paris cedex 15, France We have purified the sequence-specific DNA-binding protein KBF2 and cloned the corresponding cDNA, which is derived from the previously described RBP-JK gene, the human homolog of the Drosophila Suppressor of Hairless [Su(H)] gene. Deletion studies of the RBP-JK and Su(H) proteins allowed us to define a DNA-binding domain conserved during evolution. Because Su(H) mutant alleles exhibit dose-sensitive interactions with Hairless {H) loss-of-function mutations, we have investigated whether the RBP-JK or Su(H) proteins directly interact with the H protein in vitro. We show here that H can inhibit the DNA binding of both Su(H) and RBP-JK through direct protein-protein interactions. Consistent with this in vitro inhibitory effect, transcriptional activation driven by Su(H) in transfected Drosophila S2 cells is inhibited by H. -

Supplementary Table 1. Differentially Expressed Transcripts Between Insulin Treated (T) and Insulin Deprived (D) Type 1 Diabetic Patients
Table Legends: Supplementary Table 1. Differentially expressed transcripts between insulin treated (T) and insulin deprived (D) type 1 diabetic patients. P values were calculated using paired t-tests, and not adjusted for multiple comparison errors. Annotations for each transcript were provided by Affymetrix NetAffx Analysis Center. Supplementary Table 2. Differentially expressed mitochondrial genes between insulin treated (T) and insulin deprived (D) type 1 diabetic patients. This EXCEL file has multiple data sheets. The data sheet “MITO GENE” lists the 68 mitochondrial genes, which were grouped into 11 functional groups in 11 data sheets. P values were calculated using paired t-tests, and not adjusted for multiple comparison errors. Annotations for each transcript were provided by Affymetrix NetAffx Analysis Center. OXPHOS- oxidative phosphorylation; TCA - tricarboxylic acid cycle. Supplementary Table 3. Differentially expressed pathways by Ingenuity Pathway Analysis (IPA). This EXCEL file has two data sheets that listed the significantly up- and down-regulated pathways in insulin-deprived vs. insulin- treated patients, respectively. P values associated with each pathway were calculated by IPA. Genes were represented by HUGO symbols. Supplementary Table 1 probe set T1 T2 T3 T4 T5 T6 T7 T8 214046_at 71.6231 78.4793 69.0236 71.6228 71.3996 61.9447 66.1693 61.9322 240081_at 145.816 133.058 155.18 126.734 137.149 130.863 157.818 127.17 227644_at 132.603 132.984 144.789 127.445 134.055 115.207 141.926 126.212 233926_at 13.1944 14.7052 12.8407 -

The Plasminogen Protein Is Associated with High Myopia As
www.nature.com/scientificreports OPEN The plasminogen protein is associated with high myopia as revealed by the iTRAQ‑based proteomic analysis of the aqueous humor Kai Wen1,3, Xianfeng Shao1,3, Yahong Li1, Yaoling Li1, Yongtao Li1, Qing Wang1, Ruihong Su1, Lujie Zhang2, Yang Cai1, Jing Sun1* & Yan Zhang1* To explore the pathogenesis of high myopia (HM) using quantitative proteomics. The aqueous humor of patients with simple nuclear cataract and nuclear cataract complicated with HM (hereinafter referred to as “C” and “HM” groups, respectively) were collected. The isobaric tags for relative and absolute quantitation (iTRAQ)‑based liquid chromatography–tandem mass spectrometry (LC– MS/MS) proteomics approach was employed to explore diferentially expressed proteins (DEPs). Bioinformatics was used to interpret the proteomic results. Furthermore, the plasminogen (PLG) protein was confrmed by enzyme‑linked immunosorbent assay (ELISA) as the candidate biomarker for HM through a receiver operating characteristic curve analysis. The study showed 32 upregulated and 26 downregulated proteins. The gene ontology analysis demonstrated that 58 DEPs corresponded to 325 biological processes, 33 cell components, and 45 molecular functional annotations. The Kyoto Encyclopedia of Genes and Genomes analysis showed that the upregulated DEPs were highly enriched in the coagulation and complement cascades, consistent with the gene set enrichment analysis. Our data suggested that some DEPs might be hallmarks of the development of HM. ELISA confrmed that the PLG expression levels were signifcantly upregulated in HM. This was a new study investigating alterations in protein levels and afected pathways in HM using iTRAQ‑based quantitative proteomics. Our study provided a comprehensive dataset on overall protein changes and shed light on its potential molecular mechanism in human HM. -
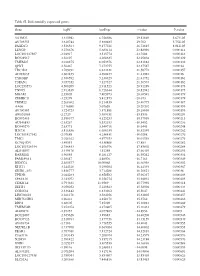
Table SI. Differentially Expressed Genes. Gene Logfc Aveexpr T Value
Table SI. Differentially expressed genes. Gene logFC AveExpr t value P‑value VSTM2L 3.315942 6.756386 29.82869 5.67E‑05 AV703555 3.610744 5.880485 29.702 5.75E‑05 FAXDC2 3.526564 9.177221 26.72815 8.01E‑05 LENG9 3.270676 5.683114 23.88996 0.000114 LOC101927507 ‑2.94917 3.515774 ‑23.7604 0.000116 BC028967 3.56197 4.208635 22.95804 0.000129 TMEM27 3.010875 8.053376 22.81882 0.000132 QPRT 2.56467 7.272755 22.37587 0.00014 TBC1D2 3.789191 6.823331 21.58772 0.000157 AF085835 2.602455 4.690477 21.43982 0.00016 TYROBP 2.386592 5.236329 21.41752 0.000161 TSPAN1 3.057352 7.127327 21.36574 0.000162 LOC253573 4.500209 2.911523 20.91289 0.000173 TNNC1 2.913839 6.726644 20.82942 0.000175 MGAM 2.35805 7.405973 20.68591 0.000179 ZDHHC11 ‑3.25279 5.412875 ‑20.673 0.000179 TRIM22 2.268602 8.214839 20.40775 0.000187 AA06 2.716698 5.07658 20.20202 0.000193 AV700385 3.724723 5.871881 20.18696 0.000193 AW851069 ‑2.2729 5.389451 ‑19.8536 0.000204 BC002465 2.590377 4.125253 19.57939 0.000213 AU146893 ‑2.63267 5.532153 ‑18.9452 0.000236 BC040270 ‑2.63771 5.540716 ‑18.6441 0.000248 H28731 3.816856 6.686159 18.32359 0.000262 LOC101927342 ‑2.07689 4.258431 ‑18.0208 0.000276 TMC1 2.028162 4.977353 18.01518 0.000276 KCNQ1DN ‑3.49551 4.538808 ‑17.883 0.000282 LOC101926934 2.786453 4.106976 17.85802 0.000284 AL043897 3.449478 4.51258 17.66439 0.000293 RARRES1 1.96055 8.281127 16.95242 0.000334 FAM189A1 ‑2.38147 4.48506 ‑16.7163 0.000349 IGDCC4 2.385577 10.00068 16.66958 0.000352 KLK11 3.244328 5.290104 16.44339 0.000367 GRID1‑AS1 1.858777 5.714288 16.26821 0.00038 UPK3B