Three New Halictine Bees from Japan (Hymenoptera, Apoidea)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
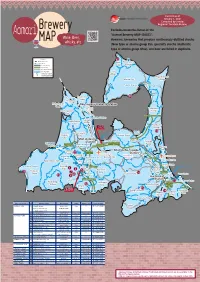
Wine, Beer, Whisky, Etc
Current as of January 1, 2021 Compiled by Sendai Regional Taxation Bureau Brewery Excludes breweries listed on the Aomori Wine, Beer, "Aomori Brewery MAP (SAKE)". MAP whisky, etc. However, breweries that produce continuously distilled shochu (New type or shochu group Ko), specialty shochu (Authentic type or shochu group Otsu), and beer are listed in duplicate. Legend ㉑ 279 Shinkansen Kazamaura JR Line Oma Private Railway Town Village Expressway Major National Highway City Boundary Sai Shinkansen Station JR and Private Railway Stations Village Higashidoori Mutsu City Village Ominato Station Shimokita Station 279 Ominato Line Imabetsu 338 ⑳ Town Sotogahama Town Nakadomari Okutsugaru-Imabetsu Station Town 338 Sotogahama Yokohama Goshogawara TsugaruTown Line Town City Kanita Station Yomogita Nakadomari Village Town Hokkaido Shinkansen Rokkasho 339 280 ①② Village 4 Noheji ⑲ Goshogawara Aomori Port Station Tsugaru City Hiranai Town City Shin-Aomori Aomori Station Noheji Town Tohoku Ajigasawa Station 101 Goshogawara Town Station Aoimori Railway394 Line Station Ou Misawa Main 103 4 Line Shichinohe-Towada City Tsuruta 338 Town ⑰ Aomori Airport Station Itayanagi Shichinohe Misawa Airport Itayanagi Town Namioka Station Aomori City Town Tohoku Shinkansen Station 394 Misawa Station Ajigasawa Town Fujisaki Kuroishi Kawabe Station Town Station 394 Oirase Town Fukaura Station Hirosaki Inakadate Hachinohe Port City ④ VillageKonan ③ Railway Kuroishi 102 Rokunohe 45 Gono Line Fukaura ⑤ City Hon- ⑯ ⑦ Hiraka 4 Town Town Towada City ⑱ Hachinohe Same Station ⑧ Station Station ⑪ ⑨ 454 Hachinohe Hirosaki Station Station ⑬ Mutsu-Minato Station Nishimeya Hirakawa Hachinohe Line Village Gonohe Town Owani⑫ City 454 ⑥⑩ Town ⑮ 7 282 Shingo Village ⑭ Hashikami Nanbu Hachinohe Town Town City Sannohe Town Name of City, Town, and Village No. -

50Must See Sites
21 Watch the sunset on Miss Veedol Beach. 36 See the Golden Line on Route 103 in the Hakkodas during the spring. 22 Visit the burial place of Christ. 37 See the fall colors at the Hirosaki Chrysanthemum Festival. 23 Check out Sakura Yabusame, an archery festival, in Towada. 38 Mountain bike through the trail at the Matsuminotaki Falls. 24 See the flower horse sculpture at the Towada City Art Museum. 39 Take in the beauty of the Kuji Sea Cliffs. 25 Check out Osorezan, also known as fear mountain. 40 See the Aomori-ken dog at the Aomori Art Musuem. 26 Have a picnic in the Statue of Liberty Park in Misawa. 41 Catch an ice hockey game in Hachinohe. 27 Collect sand dollars at Sand Dollar Beach. 42 Ride the Tsugaru Tetsudo Stove Train. 28 Go horseback riding in Towada. 43 Participate in a tea ceremony. 29 See the stalagmites in the Ryusendo Caves. 44 Enjoy a day at the beach at Tanesashi in Hachinohe. 30 Go fishing in the Pacific Ocean. 45 Ski or snowboard the 5 kilometer ski run at Appi Resort. must see sites 31 Go scuba diving off Cape Tappi. 46 Take a nice, easy walk along the Tsuta Nature Trail in the Hakkodas. 32 Sled down the astro turf slide in the Misawa Citizens Park. 47 Ride the go carts at Tonami Clan Memorial Tourist Village. 50 33 Shop for kimonos at a second hand store. 48 Take pictures of Miroku Falls near Takko-Machi. near 34 See the beautiful Juniko Lakes in Fukaura. -

A Longitudinal Analysis on the Prevalence of Overweight and Underweight Students in Aomori Prefecture in Relation to Gender, Birth Year and District
Original Article A Longitudinal Analysis on the Prevalence of Overweight and Underweight Students in Aomori Prefecture in Relation to Gender, Birth Year and District Takako Kumagai*, Ryoko Tanikawa** and Masashi Yamada** *Aomori University of Health and Welfare, Faculty of Health Sciences, Department of Nutrition 58-1 Mase, Hamadate, Aomori-shi, Aomori 030-8505 Japan [email protected] **Aomori University of Health and Welfare, Faculty of Health Sciences, Department of Nursing [Received December 7, 2018 ; Accepted July 14, 2019] Background: It has been reported that there is a pattern of onset of overweight and underweight in children and students. The prevalence of overweight children and students in Aomori Prefecture is higher than the national average for all grades, but a large-scale longitudinal analysis of overweight and underweight among children and students has not been conducted. Therefore, it has not been clarified whether there is a particular pattern in the onset of overweight or underweight. Thus, in Aomori Prefecture, large-scale longitudinal analysis on overweight and underweight of children and students is required. As the harmful effects of childhood obesity have become widely known and enlightenment activities have been carried out, overweight prevalence of children and students has been decreasing in recent years. Further reduction of overweight and underweight prevalence requires a high-risk approach, for which longitudinal data on overweight and underweight transitions is strongly required. Objective: We aim to clarify whether there is a particular pattern in the prevalence of overweight and underweight children and students in Aomori Prefecture. Methods: We constructed a pseudo-cohort using data from the Aomori Prefecture children and Student’s Health and Physical Fitness Survey Report from fiscal year (FY) 2007 to FY2018 (full survey) and the Annual Report of School Health Statistics Researches by the Ministry of Education, Culture, Sports, Science and Technology (sample survey). -

See! Eat! This Is the See! Eat! This Is
ENGLISH Enjoy Nakadomari with Your Five Senses Nakadomari is made up of the Kodomari area on the Japan Sea coast and the Nakasato area stretching inland from the Jusanko lakeside separated by Goshogawara City. The Nakadomari e region is divided in two areas that give Nakadomari both the bounty of the land and id u blessing of the sea with beautiful stereoscopic scenery unique to an area with an enclave. G g in Please visit the stunning four seasons of Nakadomari to enjoy the sights, sounds, food and e se Nakadomari ht more with all five senses. ig S et & Gourm Shin-Hakodate-Hokuto Station Aomori Spot to Eat Nakadomari Mebaru Dining Quick Look at Nakadomari Prefecture Mutsu Take a tour with Nakadomari in North Tsugaru District of Aomori Kinta Nakadomari Tappizaki Prefecture was born when Nakasato Town and and Mamejo from Kodomari Village merged into one in 2005. The new Goshogawara Aomori the Tsugaru Traditional Seikan-Tunnel-Museum Tappi Wind Park town created after this union is positioned with the Hirosaki Aomori Prefecture Kodomari Area on the Japan Sea coast and the Kinta Mamejo Puppet Hachinohe Nakasato Area located almost centrally upon the Tsugaru Peninsula away from the Kodomari area, Theater! which gives Nakadomari the sea, mountains and even a lake where people can enjoy both sightseeing Cyoukandai Observatory Seikan Tunnel and food. This is a leisurely place with a population Tatsudomari Line Hokkaido Shinkansen of roughly 12,000 (as of March 2017) where the Tsugaru Strait main industries are agricultural and fishing. Another wonderful feature is the plentiful tourist attractions Nakadomari Tsugaru-Hamana that include picturesque spots, such as the sunset Nakadomari Station melting into the Japan Sea Gongenzaki, the Tsugaru Minmaya Station Imabetsu Station Railway, lands loved by Japanese author Osamu MAP Nanatsu Sotogahama Dazai as well as the Tsugaru Traditional Kinta Falls Town Mamejo Puppet Theater, which has been designated Okawadai Station an intangible folk cultural property. -

How Garlic Saved Community? -The Efforts of JA Women's Group
How Garlic saved Community? -The efforts of JA Women's group- Ms. Fusa Sano Japan Agriculture Cooperative Hachinohe, Aomori – Overview of Hachinohe Agricultural Cooperative – President: Hiroshi Mukaiyachi Members: 17,180 people Eastern and Central Staff: 422 people Capital: 3.86 billion yen Agricultural Agricultural Gonohe- Management Center Management HQ machi Yam Strawberry Gonohe Western Hachinohe- Agricultural Management Cucumber shi Center Potato Apple Burdock Pepper root Shingo-mura Green Milk Mini-tomato Green Cucumber Beef onion Peach onion Garlic cattle Chrysanthemum Garlic Garlic Strawberry Green Spinach Green Radish beans Milk Pear Cherry onion Edible Blueberry Brewer’s Plum Tomato Chrysanthemum Hashikami- rice Rose cho Chrysanthemum Apple Calf Pepper Tomato Edamame Nanbu-cho Garlic Sannohe- Mabechi Agricultural Nanbu Agricultural machi Management Center Management Center Beef cattle Takko Agricultural Management Center Takko-machi My Hometown “Takko-machi, Aomori Prefecture” Garlic field in Ueno District shortly before harvesting season – Overview of Takko-machi, Aomori Prefecture – Population: 6,706 people (As of end of July 2010) Staples: Takko garlic, Takko beef Located in the southernmost area of Aomori Prefecture, garlic contributes Mutsu very much a part of the town’s development. Aomori Misawa Hachinohe Hirosaki Takko-machi Ninohe Kuji Akita Morioka – Local Specialties of Takko-machi, Aomori Prefecture – [Takko garlic] Takko garlic is cultivated in Takko- machi and consists of six large cloves. The garlic has a high sugar content, rich flavor, and is safe. [Takko beef] Takko cows are reared carefully in a spacious open-air area and fed with spring water. The beef is juicy and tender. [Garlic and Beef Festival] A major annual event held in the fall. -

Short Communication Nem1004 Some Additional Bionomic Characters of Deladenus Nitobei
Short communication Short communication Nem1004 Some additional bionomic characters of Deladenus nitobei Natsumi KANZAKI1,§,*, Suguru E. TANAKA2,§, Masaaki ITO3, Koji TANAKA3, Bernard SLIPPERS4 and Masanobu TABATA5 ______________________________ 1 Kansai Research Center, Forestry and Forest Products Research Institute, 68 Nagaikyutaroh, Fushimi, Kyoto 612-0855 Japan 2 Laboratory of Forest Botany, Graduate School of Agricultural and Life Sciences, The University of Tokyo, 1-1-1 Yayoi, Bunkyo-ku, Tokyo 113-8685, Japan 3 Forestry and Forest Products Research Institute, Aomori Prefectural Industrial Technology Research Center, 46-56 Kominato-Shinmichi, Hiranai, Higashi-Tsugaru, Aomori 039-3321, Japan 4 Department of Genetics, Forestry and Agricultural Biotechnology Institute (FABI), University of Pretoria, Pretoria 0002, South Africa 5 Tohoku Research Center, Forestry and Forest Products Research Institute, 92-25 Nabeyashiki, Shimo-kuriyagawa, Morioka, Iwate 020-0123, Japan § Contributed equally * Corresponding author, e-mail: [email protected] Received: 2018; revised: 2018 Accepted for publication: 2018 Keywords: emergence, parasitism, pathogenicity, Sirex nitobei. 1 Short communication Deladenus nitobei Kanzaki, Tanaka, Fitza, Kosaka, Slippers, Kimura, Tsuchiya & Tabata, 2016 was originally described as an internal parasite of a woodwasp species, Sirex nitobei Matsumura, that emerged from dead logs of Japanese red pine, Pinus densiflora Sieb. & Zucc. collected from Takko, Aomori, Japan (Kanzaki et al., 2016). During the collection of material for the species description, the emergence date and sex of the wasps were recorded, and the dissected wasp bodies were fixed in 100% ethanol and vouchered at the Forest Pathology Laboratory of Forestry and Forest Products Research Institute, Tsukuba, Japan. We re-examined the vouchered materials of the wasps and measured their body size, comparing it with the presence/absence of nematode parasitism to provide additional information about the relevance of parasitism in relation to the emergence season of the wasps. -

Life and Economy in Tsugaru, Japan. Ann D
Louisiana State University LSU Digital Commons LSU Historical Dissertations and Theses Graduate School 1999 Apples and Regional Change: Life and Economy in Tsugaru, Japan. Ann D. Brucklacher Louisiana State University and Agricultural & Mechanical College Follow this and additional works at: https://digitalcommons.lsu.edu/gradschool_disstheses Recommended Citation Brucklacher, Ann D., "Apples and Regional Change: Life and Economy in Tsugaru, Japan." (1999). LSU Historical Dissertations and Theses. 6978. https://digitalcommons.lsu.edu/gradschool_disstheses/6978 This Dissertation is brought to you for free and open access by the Graduate School at LSU Digital Commons. It has been accepted for inclusion in LSU Historical Dissertations and Theses by an authorized administrator of LSU Digital Commons. For more information, please contact [email protected]. INFORMATION TO USERS This manuscript has been reproduced from the microfilm master. UMI films the text directly from the original or copy submitted. Thus, some thesis and dissertation copies are in typewriter face, while others may be from any type of computer printer. The quality of this reproduction is dependent upon the quality of the copy subm itted. Broken or indistinct print, colored or poor quality illustrations and photographs, print bleedthrough, substandard margins, and improper alignment can adversely affect reproduction. in the unlikely event that the author did not send UMI a complete manuscript and there are missing pages, these will be noted. Also, if unauthorized copyright material had to be removed, a note will indicate the deletion. Oversize materials (e.g., maps, drawings, charts) are reproduced by sectioning the original, beginning at the upper left-hand comer and continuing from left to right in equal sections with small overlaps. -

Towada-Hachimantai National Park Page 1/ 4
TOWADA-HACHIMANTAI NATIONAL PARK PAGE 1/ 4 PG-201 TOWADA- HACHIMANTAI NATIONAL PARK This guide covers only a few of the tourist lures of northern Japan, or “Tohoku” as it’s called in Japanese, namely, the Towada- Hachimantai National Park shared by Aomori, Akita and Iwate Prefectures. This park, 85,409 ha. in area, is made up of 2 separate areas of Lake Towada and the Hachimantai Plateau, touching on the borders of Akita, Aomori and Iwate Prefectures. This district is blessed with natural wonders, which have been left unmanned to this day. Abundant hot springs are natural JR Ominato Line JR 大湊線 blessings of the district, forming Aomori 青森 Pacific Tohoku Expwy. Noheji Aomori Airport Mts.Mts. Hakkoda Hakkoda Ocean relaxing hot-spring spas here and Japan Railways(JR) 394 青森空港 Misawa Other Railways Sukayu Spa there. Most of them are small and 三沢 National Roads 酸ヶ湯温泉 Kuroishi Towadako unsophisticated, giving you a chance Excursion Boat Oirase Stream 奥入瀬渓流 Spas Hot-spring Spas Kawabe 102 Hachinohe to meet local people and experi- Hirosaki Tourist Information 八戸 ence real country life. A bus net- 弘前 Lake Towada Owani 十和田湖 work covers all the tourist spots, N Onsen but service is not very frequent and 大鰐温泉 103 104 in some areas it is suspended during 101 Towada-Minami Odate Kazuno- 282 4 the wintertime due to heavy snow 大館 Hanawa Takanosu 341 Hachimantai Plateau Noshiro 八幡平 and a cold climate. Also bus sched- Cape Toroko Spa Mt. Hachimantai Nyudozaki 7 Goshogake Spa Hachimantai ules often change with the season. Oga Spa Toshichi Spa Aspite Line Tamagawa Spa It is good to remember this if you Mt. -
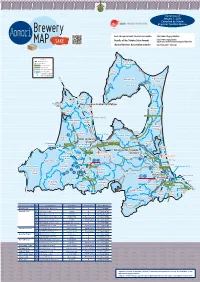
Brewery MAP SAKE
Current as of January 1, 2021 Compiled by Sendai Brewery Regional Taxation Bureau Aomori Results of the Japan Sake Awards - National New Sake Competition … https://www.nrib.go.jp/data/kan/ https://www.nta.go.jp/about/ SAKE Results of the Tohoku Sake Awards …… organization/sendai/release/kampyokai/index.htm MAP http://www.aomori-sake.or.jp/ Aomori Brewers Association website … Legend Oma 279 Shinkansen Kazamaura JR Line Town Private Railway Village Expressway Major National Highway City Boundary Sai Shinkansen Station JR and Private Railway Stations Village Mutsu City Higashidoori Village Ominato Station Shimokita Station 279 Ominato Line Imabetsu 338 Town Sotogahama Town Nakadomari Okutsugaru-Imabetsu Station Town 338 Sotogahama Yokohama Goshogawara TsugaruTown Line Town City Kanita Station Nakadomari Yomogita Town Village Hokkaido 339 Shinkansen 280 Rokkasho 4 Village Noheji Goshogawara Aomori Port Station City Hiranai Town Tsugaru City Shin-Aomori Aomori Station Noheji Town Tohoku Ajigasawa Station 101 Goshogawara Town Station Aoimori Railway394 Line Station Ou Misawa Main 103 4 Line Shichinohe-Towada City Tsuruta 338 Town Aomori Airport Station Itayanagi Namioka Station Shichinohe Misawa Airport Itayanagi Town Aomori City Town Tohoku Shinkansen Station 394 Misawa Station Ajigasawa Town Fujisaki Kuroishi Kawabe Station Town Station 394 Oirase Town Fukaura Station Hirosaki Inakadate Hachinohe Port City Village Konan Railway Kuroishi 102 Rokunohe Gono Line Fukaura City 4 Town Town Hiraka Towada City 45 Station Hachinohe Same Station Hirosaki 454 Station Station Mutsu-Minato Station Nishimeya Hirakawa Hachinohe Line Hon- Village Owani City Gonohe Town Hachinohe Town 454 Station 7 282 Shingo Village Hashikami Nanbu Hachinohe Town Town City Sannohe Town 104 Name of City, Town, and Village No. -

Open Source Used in Partner Portal Webex - Web 1.4.0
Open Source Used In Partner Portal Webex - Web 1.4.0 Cisco Systems, Inc. www.cisco.com Cisco has more than 200 offices worldwide. Addresses, phone numbers, and fax numbers are listed on the Cisco website at www.cisco.com/go/offices. Text Part Number: 78EE117C99-1101846218 Open Source Used In Partner Portal Webex - Web 1.4.0 1 This document contains licenses and notices for open source software used in this product. With respect to the free/open source software listed in this document, if you have any questions or wish to receive a copy of any source code to which you may be entitled under the applicable free/open source license(s) (such as the GNU Lesser/General Public License), please contact us at [email protected]. In your requests please include the following reference number 78EE117C99-1101846218 Contents 1.1 effective-tld-names 1.1.1 1.1.1 Available under license 1.2 brotli 1.0.9 1.2.1 Available under license 1.3 zlib 1.2.11 r3 1.3.1 Available under license 1.4 alpine-baselayout 3.2.0-r3 1.4.1 Available under license 1.5 cpp 9.3.0-r4 1.5.1 Available under license 1.6 tre 0.8.0 1.6.1 Available under license 1.7 ca-certificates 20191127-r1 1.7.1 Available under license 1.8 openssl 1.1.1i 1.8.1 Available under license 1.9 musl 1.1.24-r4 1.9.1 Available under license 1.10 icu 67 1.10.1 Available under license 1.11 node.js 15.3.0 1.11.1 Available under license 1.12 apk-tools 2.10.5-r0 1.12.1 Available under license 1.13 libuv 1.40.0 1.13.1 Available under license Open Source Used In Partner Portal Webex - Web 1.4.0 -

FACING GLOBALISATION: JAPANESE FARMERS' RESPONSES to CHANGING MARKETS Ann D. Brucklacher
FACING GLOBALISATION: JAPANESE FARMERS’ RESPONSES TO CHANGING MARKETS Ann D. Brucklacher Abstract: Internationalization has brought changes to the types of food eaten by Japanese people, and changes too to the types of produce grown by Japanese farm- ers. This paper examines changes in Japanese agriculture, looking at the various ways in which farmers are responding to and negotiating with the pressures of in- ternational and domestic food demands. Some farmers have turned to growing specialty crops, various agricultural goods with name value and organic produce, while others are experimenting with direct marketing and agricultural tourism. I focus in particular on the rise of specialty regional foods, with a description of var- ious marketing and promotion practices. 1. INTRODUCTION You are what you eat, goes the well-worn maxim of mothers and dieti- tians. And of course, our bodies do depend on the vitamins, minerals, and energy gained from food. However, beyond the pure nutritional ne- cessity of eating, we are also what we eat in terms of how we produce, prepare, market, and buy our food. Food is culture, whether manifested in the glutinous rice cakes of a Japanese New Year celebration, the wine and wafers of a Catholic ceremony, or the everyday foods we choose for comfort or as quick snacks. In Japan, however, more and more foreign foods fill the supermarket shelves, and the traditional staple rice has even been dropped from some school menus. In the age of the World Trade Organization, still more concessions to a world of free agricultural markets can be expected. Packaged foods compete with once standard fresh fruits and vegetables. -

Tohoku Brewery MAP(SAKE)(PDF/5839KB)
Current as of January 1, 2021 Compiled by Sendai Regional Taxation Bureau Click here for the results of the various awards [Japan Sake Awards - National [Tohoku Sake Awards] Brewery New Sake Competition] https://www.nrib.go.jp/ https://www.nta.go.jp/about/organization/ Tohoku data/kan/ sendai/release/kampyokai/index.htm Aomori Prefecture Aomori City ●① NISHIDA SHUZOTEN CO.,LTD. [DENSHU] SAKE Rokka Shuzo Co.,Ltd. [Joppari] Hirosaki City ●② Oma MAP Town Kazamaura ●③ MIURA SHUZO CO., LTD. [HOUHAI] Village Legend ●④ KANETA TAMADA SHUZOUTEN CO.,LTD. [HANAIPPU] Sai Matsumidori Sake Brewery Co.,Ltd. [Rokkon] Village ●⑤ Shinkansen ●⑥ MARUTAKESHUZOTEN CO.,LTD. [SHIRAKAMI ROMAN NO UTAGE] Higashidoori For information on brewery tours, please contact Mutsu City Village JR Line ●⑦ SHIRAKAMISHUZO CO.,LTD. [SHIRAKAMI] Shimokita Station Substitute Bus Line Hachinohe City ●⑧ HACHINOHE SHUZO Co.,Ltd. [Mutsu Hassen] ●⑨ HACHINOHE SHURUI CO.,LTD. Ominato Line Private Railway each site directly. HACHITSURU FACTORY [HACHITSURU] Expressway Imabetsu KIKUKOMASHUZO CO.,LTD. [Kikukoma] Town Major National Highway Gonohe Town ●⑩ Click on the name of the prefecture (read the 2D code) Sotogahama Okutsugaru-Imabetsu Station Nakadomari Town ●⑪ Hachinohe Shurui Corporation Gonohe Factory [Jyokû] Town City Boundary Sotogahama NAKAMURA KAMEKICHI CO.,LTD. [TAMADARE] Goshogawara Town Yokohama Kuroishi City ●⑫ to see an enlarged version with the phone numbers City Town Shinkansen Station Hokkaido Shinkansen ●⑬ NARUMIJOUZOUTEN CO.,LTD. [KIKUNOI] JR and Private Railway Stations Yomogita Ajigasawa Town ●⑭ OZAKI SHUZO Corporation [andou suigun] and whether brewery tours are available. Nakadomari Village Aomori Town Tsugaru Line Tsugaru City ●⑮ Takenami Shuzoten Co.,LTD. [Iwakimasamune] Port Noheji Town Rokkasho Hatomasamune Co.,LTD. [Hakkodaoroshi] Village Towada City ●⑯ 4 Shichinohe Town ●⑰ Morita Shoube Co.,Ltd.