Ginkgoales) from the Lower Cretaceous of the Iberian Ranges (Spain
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Nomenclatural Notes on Some Ginkgoalean Fossil Plants from China
植 物 分 类 学 报 45 (6): 880–883(2007) doi:10.1360/aps07015 Acta Phytotaxonomica Sinica http://www.plantsystematics.com Nomenclatural notes on some ginkgoalean fossil plants from China WU Xiang-Wu ZHOU Zhi-Yan* WANG Yong-Dong (Nanjing Institute of Geology and Palaeontology, the Chinese Academy of Sciences, Nanjing 210008, China) Abstract Two morphotaxa of ginkgoalean fossil plants from China bear illegitimate specific names, viz. Sphenobaiera biloba S. N. Feng (1977) and S. rugata Z. Q. Wang (Dec. 1984), that are heterotypic later homonyms of S. biloba Prynada (1938) and S.? rugata Z. Y. Zhou (Mar. 1984) respectively. Under Art. 53.1 and 7.3 of the Vienna Code, new specific names are proposed to supersede these illegitimate names. Two other names, viz. Baiera ziguiensis F. S. Meng (1987) and Ginkgoites elegans S. Yang, B. N. Sun & G. L. Shen (1988), were not validly published, because no nomenclatural type was definitely indicated. New species are instituted for the two morphotaxa here. Although the specific name Ginkgoites elegans Cao (1992) was antedated by Ginkgoites elegans S. Yang, B. N. Sun & G. L. Shen (1988), it still remains available for use, as the latter name has no status under the Vienna Code (Art. 12, 37) and thus no priority over Ginkgoites elegans Z. Y. Cao. Key words Ginkgoales, Ginkgoites, Baiera, Sphenobaiera, morphospecies, nomenclature. In compiling the Chinese record of fossil plants described since 1865, several illegitimate and/or not validly published names of ginkgoalean morphotaxa were found. They are either later homonyms or were published without a definite indication of the type (Art. -

Ginkgoites Ticoensis</Italic>
Leaf Cuticle Anatomy and the Ultrastructure of Ginkgoites ticoensis Archang. from the Aptian of Patagonia Author(s): Georgina M. Del Fueyo, Gaëtan Guignard, Liliana Villar de Seoane, and Sergio Archangelsky Reviewed work(s): Source: International Journal of Plant Sciences, Vol. 174, No. 3, Special Issue: Conceptual Advances in Fossil Plant Biology Edited by Gar Rothwell and Ruth Stockey (March/April 2013), pp. 406-424 Published by: The University of Chicago Press Stable URL: http://www.jstor.org/stable/10.1086/668221 . Accessed: 15/03/2013 17:43 Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at . http://www.jstor.org/page/info/about/policies/terms.jsp . JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. The University of Chicago Press is collaborating with JSTOR to digitize, preserve and extend access to International Journal of Plant Sciences. http://www.jstor.org This content downloaded on Fri, 15 Mar 2013 17:43:56 PM All use subject to JSTOR Terms and Conditions Int. J. Plant Sci. 174(3):406–424. 2013. Ó 2013 by The University of Chicago. All rights reserved. 1058-5893/2013/17403-0013$15.00 DOI: 10.1086/668221 LEAF CUTICLE ANATOMY AND THE ULTRASTRUCTURE OF GINKGOITES TICOENSIS ARCHANG. FROM THE APTIAN OF PATAGONIA Georgina M. Del Fueyo,1,* Gae¨tan Guignard,y Liliana Villar de Seoane,* and Sergio Archangelsky* *Divisio´n Paleobota´nica, Museo Argentino de Ciencias Naturales ‘‘Bernardino Rivadavia,’’ CONICET, Av. -

La Paleoflora Triásica Del Cerro Cacheuta, Provincia De Mendoza, Argentina
AMEGHINIANA - 2011 - Tomo 48 (4): 520 – 540 ISSN 0002-7014 LA PALEOFLORA TRIÁSICA DEL CERRO CACHEUTA, PROVINCIA DE MENDOZA, ARGENTINA. PETRIELLALES, CYCADALES, GINKGOALES, VOLTZIALES, CONIFERALES, GNETALES Y GIMNOSPERMAS INCERTAE SEDIS EDUARDO M. MOREL1, 2, ANALÍA E. ARTABE1, 3, DANIEL G. GANUZA1 y ADOLFO ZÚÑIGA1 1División Paleobotánica, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata. Paseo del Bosque s/n, B1900FWA La Plata, Argentina. emorel@museo. fcnym.unlp.edu.ar, [email protected], [email protected], [email protected] 2Comisión de Investigaciones Científicas de la Provincia de Buenos Aires (CIC) 3Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) Resumen. En el Cerro Cacheuta (noroeste de la provincia de Mendoza, Argentina) se relevaron cuatro perfiles de detalle, y en las localidades de Puesto Míguez y Agua de las Avispas se reconocieron siete estratos con plantas fósiles. En este aporte se presenta el estudio sistemático de las plantas fósiles encontradas y se analizan los taxones correspondientes a las Gymnospermopsida: Petriellales, Cycadales, Ginkgoales, Voltziales, Coniferales, Gnetales y Gymnospermophyta incertae sedis. El estudio sistemático incluye 25 taxones identificados como Rochipteris truncata (Frenguelli) comb. nov., Nilssonia taeniopteroides Halle, Kurtziana brandmayri Frenguelli, K. cacheutensis (Kurtz) Frenguelli, Pseudoctenis fal- coneriana (Morris) Bonetti, P. spectabilis Harris, Baiera cuyana Frenguelli, B. rollerii Frenguelli, Ginkgoidium bifidum Frenguelli, Sphenobaiera argentinae (Kurtz) Frenguelli, Heidiphyllum elongatum (Morris) Retallack, Telemachus elongatus Anderson, T. lignosus Retallack, Rissikia me- dia (Tenison-Woods) Townrow, Cordaicarpus sp., Gontriglossa sp., Yabeiella brackebuschiana (Kurtz) Ôishi, Y. mareyesiaca (Geinitz) Ôishi, Y. spathulata Ôishi, Y. wielandi Ôishi, Fraxinopsis andium (Frenguelli) Anderson y Anderson, F. -

Leppe & Philippe Moisan
CYCADALES Y CYCADEOIDALES DEL TRIÁSICO DELRevista BIOBÍO Chilena de Historia Natural475 76: 475-484, 2003 76: ¿¿-??, 2003 Nuevos registros de Cycadales y Cycadeoidales del Triásico superior del río Biobío, Chile New records of Upper Triassic Cycadales and Cycadeoidales of Biobío river, Chile MARCELO LEPPE & PHILIPPE MOISAN Departamento de Botánica, Facultad de Ciencias Naturales y Oceanográficas, Universidad de Concepción, Casilla 160-C, Concepción, Chile; e-mail: [email protected] RESUMEN Se entrega un aporte al conocimiento de las Cycadales y Cycadeoidales presentes en los sedimentos del Triásico superior de la Formación Santa Juana (Cárnico-Rético) de la Región del Biobío, Chile. Los grupos están representados por las especies Pseudoctenis longipinnata Anderson & Anderson, Pseudoctenis spatulata Du Toit y Pterophyllum azcaratei Herbst & Troncoso, y se propone una nueva especie Pseudoctenis truncata nov. sp. Las especies se encuentran junto a otros elementos típicos de las asociaciones paleoflorísticas del borde suroccidental del Gondwana. Palabras clave: Triásico superior, paleobotánica, Cycadales, Cycadeoidales. ABSTRACT A contribution to the knowledge of Cycadales and Cycadeoidales present in the Upper Triassic of the Santa Juana Formation (Carnian-Raetian) in the Bio-Bío Region of Chile is provided. The groups are represented by the species Pseudoctenis longipinnata Anderson & Anderson, Pseudoctenis spatulata Du Toit and Pterophyllum azcaratei Herbst & Troncoso. A new species Pseudoctenis truncata nov. sp. is described. They appear to be related to other typical elements of the paleofloristic assamblages from the south-occidental border of Gondwanaland. Key words: Upper Triassic, paleobotany, Cycadales, Cycadeoidales. INTRODUCCIÓN vasculares (Stewart & Rothwell 1993, Taylor & Taylor 1993). Sin embargo, se reconoce la exis- Frecuentemente se ha tratado a las Cycadeoida- tencia de una brecha morfológica entre ambos les (Bennettitales) y a las Cycadales como parte grupos, ya que a diferencia de las Medullosa- de las Cycadophyta. -

And Ginkgo Biloba Linn. Leaves
Comparative study between Ginkgoites tigrensis Archangel~ky and Ginkgo biloba Linn. leaves Liliana Villar de Seoane Liliana Villar de Seoane 1997. Comparative study between Ginkgoites tigrensis Archangelsky and GinA!go biIoba Linn. leaves. Palaeobotanist46(3) : 1-12. ' . A comparative study between leaves of Ginkgoites rigrrmsis Archangelsky'1965, which belong to the familf K.ark.e,ni:aceae and those of Ginkgo biloba L. included within the family Ginkgoaceae is realized, using lighI microscopy (LM) and scanning and transmission electron microscopy (SEM and TEM). The fossil cuticles occur in the sediments located in Baquero Formation (Lower Cretaceous), Santa Cruz Province, Argentina. The morphological, anatomical and ultrastructural analyses indicate great similarities between fossil and extant leaves. Key-words----Ginkgoalean leaves, Baquer6 Formation, Lower Cretaceous, Santa Cruz Province, Argentina. Liliana Villar de Seoane, CONICEr, Divisi6n Paleobotanica, Museo Argentino de Ciencias Naturales "B. Rivadavia ". Av. A. Gallardo 470, (1405) Buenos Aires, Argentina. RESUMEN En el presente trabajose realiza el estudio comparado entre hojas de la especie f6sil Ginkgoites tigrrmsis Archangelsky 1965, pel1eneciente a la familia Karkeniaceae, y de la especie actual Ginkgo biloba L. pel1eneciente a la familia Ginkgoaceae, ambas familias incluidas dentro del orden Ginkgoales. Para efectuar el anAlisis cuticularse utiliz6 microscopia 6ptica (MO) y microscopia electronica de barrido y transmision (MEB y MEn. Los reslOS f6siles analizados fueron extraidos de sedimentitas organ6genas hlladas en la Formacion Baquer6 de la Provincia de Santa Cruz (Cret:!icico Inferior). Los resultados obtenidos con los anAlisis realizados, han demostrado la existencia de grandes semejanzas morfologicas, anat6micas y ultraestructurales entre las hojas de las especies Ginkgoites tigrrmsis y Ginkgo biloba. -

La Paleoflora Triásica De Potrerillos, Provincia De Mendoza, Argentina
AMEGHINIANA (Rev. Asoc. Paleontol. Argent.) - 44 (2): 279-301. Buenos Aires, 30-6-2007 ISSN 0002-7014 La paleoflora triásica de Potrerillos, provincia de Mendoza, Argentina Analía E. ARTABE1,2, Eduardo M. MOREL1,3 Daniel G. GANUZA1, Ana M. ZAVATTIERI4 y Luis A. SPALLETTI5 Abstract. THE TRIASSIC PALEOFLORA OF POTRERILLOS, MENDOZA PROVINCE, ARGENTINA. At the Bayo and Cocodrilo Hills (Potrerillos, northwestern Mendoza Province, Argentina) two stratigraphic logs were measured in detail and 16 plantiferous levels were determined. In this contribution the systematic study of the paleoflora is presented. The 23 taxa recognized belong to Sphenopsida (Equisetales), Filicopsida (Filicales) and Gymnospermopsida (Corystospermales, Peltaspermales, Cycadales, Ginkgoales, Petriellales and Gnetales). The following taxa are des- cribed and compared: Equisetites fertilis (Frenguelli) Frenguelli, Neocalamites carrerei (Zeiller) Halle, Cladophlebis me- sozoica Kurtz ex Frenguelli, Cladophlebis mendozaensis (Geinitz) Frenguelli, Dicroidium argenteum (Retallack) Gnae- dinger and Herbst, Dicroidium dubium (Feistmantel) Gothan, Dicroidium odontopteroides (Morris) Gothan, Johnstonia stelzneriana (Geinitz) Frenguelli, Johnstonia coriacea (Johnston) Walkom, Xylopteris elongata (Carruthers) Frenguelli, Zuberia feistmanteli (Johnston) Frenguelli, Feruglioa samaroides Frenguelli, Pachydermophyllum praecordillerae (Fren- guelli) Retallack, Kurtziana cacheutensis (Kurtz) Frenguelli emend. Petriella and Arrondo, Sphenobaiera argentinae (Kurtz) Frenguelli, Baiera cuyana -

An Unexpected Noncarpellate Epigynous Flower from the Jurassic
RESEARCH ARTICLE An unexpected noncarpellate epigynous flower from the Jurassic of China Qiang Fu1, Jose Bienvenido Diez2, Mike Pole3, Manuel Garcı´aA´ vila2,4, Zhong-Jian Liu5*, Hang Chu6, Yemao Hou7, Pengfei Yin7, Guo-Qiang Zhang5, Kaihe Du8, Xin Wang1* 1CAS Key Laboratory of Economic Stratigraphy and Paleogeography, Nanjing Institute of Geology and Palaeontology and Center for Excellence in Life and Paleoenvironment, Chinese Academy of Sciences, Nanjing, China; 2Departamento de Geociencias, Universidad de Vigo, Vigo, Spain; 3Queensland Herbarium, Brisbane Botanical Gardens Mt Coot-tha, Toowong, Australia; 4Facultade de Bioloxı´a, Asociacio´n Paleontolo´xica Galega, Universidade de Vigo, Vigo, Spain; 5State Forestry Administration Key Laboratory of Orchid Conservation and Utilization at College of Landscape Architecture, Fujian Agriculture and Forestry University, Fuzhou, China; 6Tianjin Center, China Geological Survey, Tianjin, China; 7Key Laboratory of Vertebrate Evolution and Human Origin of Chinese Academy of Sciences, Institute of Vertebrate Paleontology and Paleoanthropology and Center for Excellence in Life and Paleoenvironment, Chinese Academy of Sciences, Beijing, China; 8Jiangsu Key Laboratory for Supramolecular Medicinal Materials and Applications, College of Life Sciences, Nanjing Normal University, Nanjing, China Abstract The origin of angiosperms has been a long-standing botanical debate. The great diversity of angiosperms in the Early Cretaceous makes the Jurassic a promising period in which to anticipate the origins of the angiosperms. Here, based on observations of 264 specimens of 198 *For correspondence: individual flowers preserved on 34 slabs in various states and orientations, from the South [email protected] (Z-JL); Xiangshan Formation (Early Jurassic) of China, we describe a fossil flower, Nanjinganthus [email protected] (XW) dendrostyla gen. -

Late Permian) Flora from the Bletterbach Gorge in the Dolomites, Northern Italy: a Review 39-61 Geo.Alp, Vol
ZOBODAT - www.zobodat.at Zoologisch-Botanische Datenbank/Zoological-Botanical Database Digitale Literatur/Digital Literature Zeitschrift/Journal: Geo.Alp Jahr/Year: 2017 Band/Volume: 0014 Autor(en)/Author(s): Kustatscher Evelyn, Konijnenburg-van Cittert Johanna H.A. van, Looy Cindy V., Labandeira Conrad, Wappler Thorsten, Butzmann Rainer, Fischer Thilo C., Krings Michael, Kerp Hans, Visscher Henk Artikel/Article: The Lopingian (late Permian) flora from the Bletterbach Gorge in the Dolomites, Northern Italy: a review 39-61 Geo.Alp, Vol. 14 2017 The Lopingian (late Permian) flora from the Bletterbach Gorge in the Dolomites, Northern Italy: a review • Evelyn Kustatscher1,2, Johanna H.A. van Konijnenburg-van Cittert3,4, Cindy V. Looy5, Conrad C. Labandeira6–8, Torsten Wappler9, Rainer Butzmann10, Thilo C. Fischer11, Michael Krings2, Hans Kerp12 & Henk Visscher4 1 Museum of Nature South Tyrol, Bindergasse/Via Bottai 1, 39100 Bozen/Bolzano, Italy; email: [email protected] 2 Department für Geo- und Umweltwissenschaften, Paläontologie und Geobiologie, Ludwig-Maximilians-Universität, and SNSB Bayerische Staatssammlung für Paläontologie und Geologie, Richard-Wagner-Straße 10, 80333 Munich, Germany; e-mail: [email protected] 3 National Centre for Biodiversity ‘Naturalis’, PO Box 9517, 2300 RA Leiden; e-mail: [email protected] 4 Laboratory of Palaeobotany and Palynology, Faculty of Geosciences, Utrecht University, Heidelberglaan 2, 3584 CS Utrecht, email: [email protected]; [email protected] 5 Department -

Flora of the Late Triassic
Chapter 13 Flora of the Late Triassic Evelyn Kustatscher, Sidney R. Ash, Eugeny Karasev, Christian Pott, Vivi Vajda, Jianxin Yu, and Stephen McLoughlin Abstract The Triassic was a time of diversification of the global floras following the mass-extinction event at the close of the Permian, with floras of low-diversity and somewhat uniform aspect in the Early Triassic developing into complex vegetation by the Late Triassic. The Earth experienced generally hothouse conditions with low equator-to-pole temperature gradients through the Late Triassic. This was also the time of peak amalgamation of the continents to form Pangea. Consequently, many plant families and genera were widely distributed in the Late Triassic. Nevertheless, E. Kustatscher (*) Museum of Nature South Tyrol, Bindergasse 1, 39100 Bozen/Bolzano, Italy Department für Geo– und Umweltwissenschaften, Paläontologie und Geobiologie, Ludwig– Maximilians–Universität, and Bayerische Staatssammlung für Paläontologie und Geologie, Richard–Wagner–Straße 10, 80333 Munich, Germany e-mail: [email protected] S.R. Ash Department of Earth and Planetary Sciences, Northrop Hall, University of New Mexico, Albuquerque, NM 87131, USA e-mail: [email protected] E. Karasev Borissiak Paleontological Institute, Russian Academy of Sciences, Profsoyuznaya 123, Moscow 117647, Russia e-mail: [email protected] C. Pott Palaeobiology Department, Swedish Museum of Natural History, P.O. Box 50007, SE-104 05 Stockholm, Sweden LWL-Museum of Natural History, Westphalian State Museum and Planetarium, Sentruper Straße 285, 48161 Münster, Germany e-mail: [email protected] V. Vajda • S. McLoughlin Palaeobiology Department, Swedish Museum of Natural History, P.O. Box 50007, SE-104 05 Stockholm, Sweden e-mail: [email protected]; [email protected] J. -
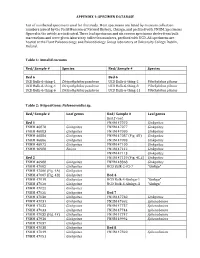
Appendix 1: Specimen Database
APPENDIX 1: SPECIMEN DATABASE List of numbered specimens used for this study. Most specimens are listed by museum collection numbers issued by the Field Museum of Natural History, Chicago, and prefixed with FNHM. Specimens figured in the article are indicated. Three leaf specimens and six cocoon specimens derive from bulk macerations and were given laboratory collection numbers, prefixed with UCD. All specimens are hosted at the Plant Palaeoecology and Palaeobiology Group laboratory at University College Dublin, Ireland. Table 1: Annelid cocoons Bed/ Sample # Species Bed/ Sample # Species Bed 6 Bed 6 UCD Bulk-6-thing-1 Dictyothylakos pesslerae UCD Bulk-6-thing-2 Pilothylakos pilosus UCD Bulk-6-thing-4 Dictyothylakos pesslerae UCD Bulk-6-thing-8 Pilothylakos pilosus UCD Bulk-6-thing-6 Dictyothylakos pesslerae UCD Bulk-6-thing-11 Pilothylakos pilosus Table 2: Ovipositions: Palaeoovoidus sp. Bed/ Sample # Leaf genus Bed/ Sample # Leaf genus Bed 2 cont. Bed 1 FNHM 47070 Ginkgoites FNHM 46870 Ginkgoites FNHM 47077 Ginkgoites FNHM 46883 Ginkgoites FNHM 47080 Ginkgoites FNHM 46884 Ginkgoites FNHM 47087 (Fig. 4D) Ginkgoites FNHM 46886 Ginkgoites FNHM 47098 Ginkgoites FNHM 46972 Ginkgoites FNHM 47100 Ginkgoites FNHM 46980 Baiera FNHM 47112 Ginkgoites FNHM 47113 Ginkgoites Bed 2 FNHM 47119 (Fig. 4C,E) Ginkgoites FNHM 46988 Ginkgoites FNHM 48868 Ginkgoites FNHM 47005 Ginkgoites UCD Bulk-2-05-7 “Ginkgo” FNHM 47006 (Fig. 4A) Ginkgoites FNHM 47007 (Fig. 4B) Ginkgoites Bed 4 FNHM 47019 Ginkgoites UCD Bulk-4-Ginkgo-1 “Ginkgo” FNHM 47020 Ginkgoites UCD Bulk-4-Ginkgo-3 “Ginkgo” FNHM 47022 Ginkgoites FNHM 47028 Ginkgoites Bed 7 FNHM 47030 Ginkgoites FNHM 47760 Ginkgoites FNHM 47031 Ginkgoites FNHM 47692 Sphenobaiera FNHM 47032 Ginkgoites FNHM 47737 Sphenobaiera FNHM 47033 Ginkgoites FNHM 47744 Sphenobaiera FNHM 47035 (Fig. -
Middle-Late Albian of Spain)
G Model GEOBIO-844; No. of Pages 7 Geobios xxx (2018) xxx–xxx Available online at ScienceDirect www.sciencedirect.com Original article New data about Early Cretaceous Ginkgoales from southwestern § Europe (middle-late Albian of Spain) a,b, c a Luis Miguel Sender *, Jose´ Bienvenido Diez , Javier Ferrer a ´ Area de Paleontologı´a, Facultad de Ciencias Edificio C (Geolo´gicas), Universidad de Zaragoza, C/Pedro Cerbuna s/n., 50009 Zaragoza, Spain b Department of Biological Sciences, Faculty of Science and Engineering, Chuo University, 1-13-27 Kasuga Bunkyo-ku, 1128551 Tokyo, Japan c Departamento de Xeociencias Marin˜as e Ordenacio´n do Territorio, Facultade de Ciencias do Mar, Universidade de Vigo, 36310 Vigo Galicia, Spain A R T I C L E I N F O A B S T R A C T Article history: We present in this work a leaf-type corresponding to the Ginkgoales genus Sphenobaiera recorded in Received 30 May 2017 deposits from the middle-upper Albian (Lower Cretaceous) of northeastern Spain. The fossil leaf shows a Accepted 4 August 2018 widely dissected lamina that forks regularly six times ending at the apex in sixty-four tiny wedge-shaped Available online xxx segments, which are characters similar to those in Sphenobaiera pecten from the Middle Jurassic of England. The exceptional preservation of this fossil leaf, which preserves all segments in connection with Keywords: its base, in addition to taphonomical and sedimentological data, indicates a nearby location for the Sphenobaiera parent plant, which grew near the shores of a tidally-influenced fluvial environment. A palaeoenvi- Taphonomy ronmental reconstruction of the sub-environments where the different types of Ginkgoales known from Tidally-influenced environments Palaeophytogeography the Albian of northeastern Spain developed is also presented. -

Revisiting the Mid-Permian Agay Flora of the Estérel Basin (Provence, South-East France)
Revisiting the mid-Permian Agay flora of the Estérel basin (Provence, south-east France) Iris Kuipers (3703584) MSc Thesis master Earth, Life and Climate, Utrecht University Abstract The conifer-dominated mid-Permian flora of Agay (Estérel Massif, Provence, south-east France) was first described by Visscher (1968). The present study revisits Visscher’s publication by collating it with new material, arriving at a total count of 282 fossils. The analysis of these fossils yielded a similar species list in which the conifer Ullmannia bronnii is by far the most abundant. Other commonly found conifers are Ullmannia frumentaria, Quadrocladus solmsii and Quadrocladus orobiformis. The remaining 8% of the assemblage, contains the seed fern Sphenopteris, the holotype of the (putative) ginkgophyte Esterella gracilis and several fossils of uncertain affinity (inter alia the holotype of Sp. A Boersma and Visscher (1969)). This species composition shows similarities to the coniferous late Permian Zechstein flora. Combining the most recent paleomagnetic data and radiometric dating with information from other Permian basins in the Provence suggests a Wordian (mid-Permian) age for the Estérel flora. Introduction The formation of the supercontinent Pangea that started during the Carboniferous was almost complete at the end of the Permian (298.9 - 252.2 Ma) (Stanley, 2005). Due to the northward movement of Gondwana and its subsequent collision with Laurasia, the landscape of Pangea was very mountainous with extended dune deposits and evaporites in its continental interior developing under strong aridity caused by the great land-sea distance. The aridity on Pangea persisted and increased throughout its formation and existence. The warming by solar radiation and heat loss by longwave back radiation of Pangea’s major land masses, located in the mid-latitudes, caused a very high seasonal contrast resulting in a strong monsoonal circulation in which winters became very cold and dry and summers became very warm and wet (Ruddiman 2001, Roscher and Schneider 2006).