The Human Gluteus Maximus and Its Role in Running
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Hip Extensor Mechanics and the Evolution of Walking and Climbing Capabilities in Humans, Apes, and Fossil Hominins
Hip extensor mechanics and the evolution of walking and climbing capabilities in humans, apes, and fossil hominins Elaine E. Kozmaa,b,1, Nicole M. Webba,b,c, William E. H. Harcourt-Smitha,b,c,d, David A. Raichlene, Kristiaan D’Aoûtf,g, Mary H. Brownh, Emma M. Finestonea,b, Stephen R. Rossh, Peter Aertsg, and Herman Pontzera,b,i,j,1 aGraduate Center, City University of New York, New York, NY 10016; bNew York Consortium in Evolutionary Primatology, New York, NY 10024; cDepartment of Anthropology, Lehman College, New York, NY 10468; dDivision of Paleontology, American Museum of Natural History, New York, NY 10024; eSchool of Anthropology, University of Arizona, Tucson, AZ 85721; fInstitute of Ageing and Chronic Disease, University of Liverpool, Liverpool L7 8TX, United Kingdom; gDepartment of Biology, University of Antwerp, 2610 Antwerp, Belgium; hLester E. Fisher Center for the Study and Conservation of Apes, Lincoln Park Zoo, Chicago, IL 60614; iDepartment of Anthropology, Hunter College, New York, NY 10065; and jDepartment of Evolutionary Anthropology, Duke University, Durham, NC 27708 Edited by Carol V. Ward, University of Missouri-Columbia, Columbia, MO, and accepted by Editorial Board Member C. O. Lovejoy March 1, 2018 (received for review September 10, 2017) The evolutionary emergence of humans’ remarkably economical their effects on climbing performance or tested whether these walking gait remains a focus of research and debate, but experi- traits constrain walking and running performance. mentally validated approaches linking locomotor -

Rehabilitation of Soft Tissue Injuries of the Hip and Pelvis T
Donald and Barbara Zucker School of Medicine Journal Articles Academic Works 2014 Rehabilitation of soft tissue injuries of the hip and pelvis T. F. Tyler Northwell Health T. Fukunaga Hofstra Northwell School of Medicine J. Gellert Follow this and additional works at: https://academicworks.medicine.hofstra.edu/articles Recommended Citation Tyler TF, Fukunaga T, Gellert J. Rehabilitation of soft tissue injuries of the hip and pelvis. 2014 Jan 01; 9(6):Article 1396 [ p.]. Available from: https://academicworks.medicine.hofstra.edu/articles/1396. Free full text article. This Article is brought to you for free and open access by Donald and Barbara Zucker School of Medicine Academic Works. It has been accepted for inclusion in Journal Articles by an authorized administrator of Donald and Barbara Zucker School of Medicine Academic Works. INVITED CLINICAL COMMENTARY REHABILITATION OF SOFT TISSUE INJURIES OF THE HIP AND PELVIS Timothy F. Tyler MS, PT, ATC1 Takumi Fukunaga DPT, ATC, CSCS1 Joshua Gellert DPT IJSPT ABSTRACT Soft tissue injuries of the hip and pelvis are common among athletes and can result in significant time loss from sports participation. Rehabilitation of athletes with injuries such as adductor strain, iliopsoas syn- drome, and gluteal tendinopathy starts with identification of known risk factors for injury and comprehen- sive evaluation of the entire kinetic chain. Complex anatomy and overlapping pathologies often make it difficult to determine the primary cause of the pain and dysfunction. The purpose of this clinical commen- tary is to present an impairment-based, stepwise progression in evaluation and treatment of several com- mon soft tissue injuries of the hip and pelvis. -

All About Glutes 1 Table of Contents
All About Glutes 1 Table of Contents Are You Training Your Glutes the Wrong Way? 3 • Anatomy of the Glutes 4 • Functions of the Glutes at the Hip 4 • The Shortcoming of Most Training Programs 5 • Progression and Preventing Knee Valgus 6 • Simple Solution 6 How to Identify and Correct Tight Hip Flexors 8 • What Exactly Are Tight Hip Flexors? 9 • The Hip Flexor Muscle Group 9 • Signs You Have Tight Hip Flexors 10 • What Causes Hip Tightness 10 • Stretches to Loosen up Tight Hip Flexors 10 • Exercises to Strengthen Hip Flexors 11 Pain in the Buttocks When Sitting? Tips to Prevent and Manage 12 Piriformis Syndrome • What is Piriformis Syndrome? 13 • How Does Piriformis Syndrome Happen? 13 • Special Considerations with Clients 14 • Prevention and Pain Management 14 How Do I Build the Perfect Glutes? 16 • Can’t I Just Squat and Lunge? 17 • Your Best Bets to Target the Glutes 18 • Don’t Forget the Legs 18 • Train the Glutes SPECIFICALLY 19 TABLE OF CONTENTS 800.545.4772 WWW.ISSAONLINE.EDU 2 Are You Training Your Glutes the Wrong Way? 800.545.4772 WWW.ISSAONLINE.EDU 3 UNIT ONE These days, the glutes get a lot of attention, and it’s well deserved. When you build and strengthen your glutes in the right way, they not only make your body look better, but they also increase your performance and can diminish knee pain. The problem is most people aren’t taking the best approach to training for the highest level of glute development. Anatomy of the Glutes Let’s start with a little anatomy. -

Importance of Iliopsoas and Erector Spinae Muscles in Predicting The
International Journal of Physiotherapy and Research, Int J Physiother Res 2014, Vol 2(5):681-88. ISSN 2321-1822 Original Article IMPORTANCE OF ILIOPSOAS AND ERECTOR SPINAE MUSCLES IN PREDICTING THE FUNCTIONAL COMPETENCE OF TRANSFEMORAL AMPUTEES Lajja K Rishi 1, Suraj Kumar *2, Sangeeta Lahiri 3, V.P. Sharma 4. 1 Master of Physiotherapy (student), National Institute for the Orthopaedically Handicapped, Kolkata, West Bengal, India. *2 HOD and Associate Professor, Department of Physiotherapy, UPRIMS&R, Saifai, Etawah, UP, India. 3 Physical Therapist (cardio-respiratory disorders & intensive care), India. 4 Professor, KGM Medical University, Lucknow, India. ABSTRACT Purpose: Muscle imbalance in transfemoral amputees impair physical mobility and activities of daily living. Aim of this study was to correlate the muscle imbalance with functional competence in transfemoral amputees. Methods: Thirty amputees were evaluated under inclusion criteria and randomly allocated into 2 groups. Group A received stretching(1 week) followed by strengthening(3 weeks) and in group B strengthening(3 weeks) were followed by stretching(1 week) . Phase I includes values after 1 week stretching program in group A and 3 weeks strengthening program in group B. Data were recorded at baseline, after phase I completion and end of treatment. Physical mobility was assessed by “Timed up and go” test. Results: Muscle imbalance and physical mobility improved significantly in both groups at the end of treatment. The correlation values of “Timed up and go” test with Iliopsoas and Erector spinae muscle showed significant improvement in both groups. Conclusion: Baseline measurements showed that Iliopsoas and Erector spinae muscles were tight whereas Gluteus maximus and Abdominal muscles were weak in transfemoral amputees. -

Effects of a Gluteal Muscles Specific Exercise Program on the Vertical
International Journal of Environmental Research and Public Health Article Effects of a Gluteal Muscles Specific Exercise Program on the Vertical Jump Tomás Gallego-Izquierdo 1, Gerardo Vidal-Aragón 2, Pedro Calderón-Corrales 2, Álvaro Acuña 2, Alexander Achalandabaso-Ochoa 3,* , Agustín Aibar-Almazán 3 , Antonio Martínez-Amat 3 and Daniel Pecos-Martín 1 1 Physiotherapy and Pain Group, Department of Physical Therapy, University of Alcala, 28801 Madrid, Spain; [email protected] (T.G.-I.); [email protected] (D.P.-M.) 2 Physical Therapist, Department of Physical Therapy, University of Alcala, 28801 Madrid, Spain; [email protected] (G.V.-A.); pedrocalderon.fi[email protected] (P.C.-C.); [email protected] (Á.A.) 3 Department of Health Sciences, Faculty of Health Sciences, University of Jaén, 23071 Jaén, Spain.; [email protected] (A.A.-A.); [email protected] (A.M.-A.) * Correspondence: [email protected]; Tel.: +34-953213651 Received: 9 June 2020; Accepted: 23 July 2020; Published: 27 July 2020 Abstract: The vertical jump is a complex movement where many factors are involved in the final result. Currently, how a specific exercise program for gluteal muscles can affect the vertical jump is unknown. So, the aim of this study was to examine the effect of a specific exercise program for the gluteal muscles on a vertical jump. Forty-nine amateur athletes completed an 8-week program. The experimental group received a specific gluteal muscle training program in addition to their regular training routine, whereas the control group received their regular training routine. Jump height, flight time, speed and power were assessed (baseline, postintervention, and 4-week follow-up). -

ACE's Essentials of Exercise Science for Fitness Professionals
ACE’s Essentials of Exercise Science for Fitness Professionals Chapter 3: Fundamentals of Applied Kinesiology Hip Flexors . There are 21 major muscles involved in the actions of the hip joint. – Actions of the hip joint include flexion, extension, internal rotation, external rotation, adduction, and abduction. – More than half of these muscles are involved in multiple actions. Anterior Hip Muscles: Hip Flexors . Active range of motion for hip flexion . Prime movers: iliopsoas, rectus femoris, sartorius, pectineus, and tensor fasciae latae – Act synergistically to cause hip flexion (e.g., “up” phase of a knee lift) – Act eccentrically to control hip extension (e.g., “down” phase of a knee lift) Hip Flexors: Considerations . Muscle origins and insertions impact muscular function. Iliopsoas – The psoas major and Sample Sample stretching exercise: minor originate in the strengthening low back and insert to the exercise: proximal femur, leading to poor mechanical leverage when used to raise and lower a straight leg. – The abdominals are not strong enough to balance the large force and keep the spine in neutral alignment. Hip Flexors: Considerations . Rectus femoris – Works at both the knee and hip, concentrically contracting to perform hip flexion and knee extension. – Sample strengthening exercise: standing straight-leg raise – Sample stretching exercise: iliopsoas lunge, bending the back knee . The sartorius is the longest muscle in the body. – It is also involved in hip abduction, adduction, and external rotation, and knee flexion and internal rotation. Tensor fascia latae (TFL) – IT band – Explosive hip flexion results in a highly developed TFL. Hip Internal and External Rotators . The hip internal rotators include the tensor fasciae latae, semitendinosus (slight), and semimembranosus (slight). -

The Absence of Piriformis Muscle, Combined Muscular Fusion, and Neurovascular Variation in the Gluteal Region
Autopsy Case Report The absence of piriformis muscle, combined muscular fusion, and neurovascular variation in the gluteal region Matheus Coelho Leal1 , João Gabriel Alexander1 , Eduardo Henrique Beber1 , Josemberg da Silva Baptista1 How to cite: Leal MC, Alexander JG, Beber EH, Baptista JS. The absence of piriformis muscle, combined muscular fusion, and neuro-vascular variation in the gluteal region. Autops Case Rep [Internet]. 2021;11:e2020239. https://doi.org/10.4322/ acr.2020.239 ABSTRACT The gluteal region contains important neurovascular and muscular structures with diverse clinical and surgical implications. This paper aims to describe and discuss the clinical importance of a unique variation involving not only the piriformis, gluteus medius, gluteus minimus, obturator internus, and superior gemellus muscles, but also the superior gluteal neurovascular bundle, and sciatic nerve. A routine dissection of a right hemipelvis and its gluteal region of a male cadaver fixed in 10% formalin was performed. During dissection, it was observed a rare presentation of the absence of the piriformis muscle, associated with a tendon fusion between gluteus and obturator internus, and a fusion between gluteus minimus and superior gemellus muscles, along with an unusual topography with the sciatic nerve, which passed through these group of fused muscles. This rare variation stands out with clinical manifestations that are not fully established. Knowing this anatomy is essential to avoid surgical iatrogeny. Keywords Anatomic Variation; Anatomy; Buttocks; Muscle; Piriformis Muscle Syndrome. INTRODUCTION The gluteal region contains important Over the years, these variations have been neurovascular and muscular structures that may classified and distributed into different groups. impose diverse clinical and surgical approaches. -
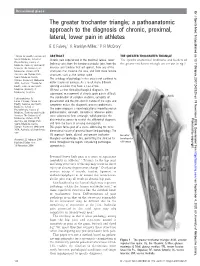
The Greater Trochanter Triangle
Occasional piece Br J Sports Med: first published as 10.1136/bjsm.2007.042325 on 19 November 2008. Downloaded from The greater trochanter triangle; a pathoanatomic approach to the diagnosis of chronic, proximal, lateral, lower pain in athletes E C Falvey,1 A Franklyn-Miller,1 P R McCrory2 1 Centre for Health, Exercise and ABSTRACT THE GREATER TROCHANTER TRIANGLE Sports Medicine, School of Chronic pain experienced in the proximal, lateral, lower The specific anatomical landmarks and borders of Physiotherapy, Faculty of Medicine, Dentistry and Health limb may arise from the femoro-acetabular joint, from the the greater trochanter triangle are set out in fig 1. Sciences The University of muscles and tendons that act upon it, from any of the Melbourne, Victoria 3010 structures that traverse the area, and from more remote Australia and Olympic Park structures such as the lumbar spine. Sports Medicine Centre, The aetiology of pathology in this area is not confined to Olympic Boulevard, Melbourne 3004, Australia; 2 Centre for either trauma or overuse. As a result many different Health, Exercise and Sports sporting activities may have a causal role. Medicine University of Without a clear clinical/pathological diagnosis, the Melbourne, Australia subsequent management of chronic groin pain is difficult. Correspondence to: The combination of complex anatomy, variability of Eanna C Falvey, Centre for presentation and the non-specific nature of the signs and Health, Exercise and Sports symptoms makes the diagnostic process problematic. Medicine, School of Physiotherapy, Faculty of The paper proposes a novel educational model based on Medicine, Dentistry and Health pathoanatomic concepts. -

Gluteal Region and Back of Thigh Doctors Notes Notes/Extra Explanation Editing File Objectives
Color Code Important Gluteal Region and Back of Thigh Doctors Notes Notes/Extra explanation Editing File Objectives Know contents of gluteal region: Groups of Glutei muscles and small muscles (Lateral Rotators). Nerves & vessels. Foramina and structures passing through them as: 1-Greater Sciatic Foramen. 2-Lesser Sciatic Foramen. Back of thigh : Hamstring muscles. Movements of the lower limb Hip = Thigh Knee=Leg Foot=Ankle Flexion/Extension Flexion/Extension Flexion/Extension Rotation Adduction/Abduction Inversion/Eversion Contents Of Gluteal Region: Muscles / Nerves / Vessels 1- Muscles: • Glutei: 1. Gluteus maximus. 2. Gluteus medius. 3. Gluteus minimus. Abductors: • Group of small muscles (Lateral Rotators): 1. Gluteus medius. 2. Gluteus minimus. 1.Piriformis. Rotators: 2.Obturator internus 1. Obturator internus. 3.Superior gemellus 2. Quadratus femoris. 4.Inferior gemellus Extensor: 5.Quadratus femoris Gluteus maximus. Contents Of Gluteal Region: Muscles / Nerves / Vessels 2- Nerves (All from Sacral Plexus): 1. Sciatic nerve. 2. Superior gluteal nerve. 3. Inferior gluteal nerve. 4. Post. cutaneous nerve of thigh. 5. Nerve to obturator internus. 6. Nerve to quadratus femoris. 7. Pudendal nerve. Contents Of Gluteal Region: Muscles / Nerves / Vessels 3- VESSELS: (all from internal iliac vessels): 1. Superior gluteal 2. Inferior gluteal 3. Internal pudendal vessels. Greater sciatic foreamen: Greater sciatic notch of hip bone is transformed into foramen by: sacrotuberous (between the sacrum to ischial tuberosity) & sacrospinous (between the sacrum to ischial spine ) Structures passing through Greater sciatic foramen : Nerves: Vessels: Greater sciatic foramen Above 1. Superior gluteal nerves, 2. Superior gluteal piriformis vessels. Lesser sciatic foramen muscle. 3. Piriformis muscle. Belew 4. Inferior gluteal nerves 10. -

Strengthening Your Hip Muscles Some Exercises May Be Better Than Others
jospt perspectives for patients Strengthening Your Hip Muscles Some Exercises May Be Better Than Others J Orthop Sports Phys Ther 2013;43(2):65. doi:10.2519/jospt.2013.0501 eak hip muscles lead to poor hip motion, and poor difficult to strengthen these muscles without also strengthening hip motion can cause knee, hip, and back pain. By a muscle called the tensor fascia lata, which is located toward exercising to strengthen the hip muscles that con- the front of the hip. Too much activation of that muscle may Wtrol how your hip moves, you can reduce your pain create unwanted hip motion that may worsen knee, hip, or back in these parts of your body. The 2 key muscles to include in your pain. A study published in the February 2013 issue of JOSPT exercise program are the gluteus maximus (the chief muscle on provides information intended to help physical therapists and the back of your hip—your buttocks) and the gluteus medius their patients select exercises that target the buttock muscles (the main muscle on the side of your hip). However, it is often without causing other unwanted muscle actions. NEW INSIGHTS In this study, the researchers had 20 healthy people perform 11 different hip exercises commonly used for both fitness and rehabilitation. While the participants performed the exercises, fine wires were used to record the amount of electrical activity within the 3 muscles. This indicated how much each muscle was working. The researchers’ goal was to discover which exercises used the gluteus maximus and gluteus medius muscles the most, while minimizing the action of the tensor fascia lata. -

Pelvis & Thigh
Pelvis & Thigh 6 After meeting a stranger, you soon begin to palpate their piriformis Topographical Views 276 muscle (located deep in the posterior buttock). You certainly wouldn’t try Exploring the Skin and Fascia 277 this in “everyday life,” but in patient care settings this level of familiarity is Bones of the Pelvis and Thigh 278 commonplace—and welcomed by a client with a hypercontracted piriformis. Bony Landmarks of the Pelvis Touch is a unique privilege afforded to health care providers. As such, we and Thigh 279 need to be mindful of the trust our clients have in us. One way to insure this Overview: Bony Landmark Trails 284 is through good communication skills. For instance, working the adductors Overview: Muscles of the and gluteal region requires a practitioner to provide ample explanation as to Pelvis and Thigh 296 the rationale, need, and goals of working these intimate areas of the body. Synergists—Muscles Working This chapter might pose new challenges for you, as we will be palpating Together 302 structures close to intimate areas. Muscles of the Pelvis and Thigh 306 Ligaments and Other Before proceeding, consider the following questions: Structures of the Pelvis and Thigh 336 E Have you ever been anxious to undergo a physical exam? Was there anything the practitioner did or could have done to alleviate this anxiety? Consider multiple elements, including both verbal and nonverbal communication, draping, physical pressure, and pace. E Tissues and landmarks found in the pelvis and thigh tend to be significantly larger than those discussed in previous chapters. How might your palpation techniques need to change? E Also, how might you properly and comfortably position your patient to access structures needing to be palpated. -

An Unusual Presentation of Right-Sided Sciatica with Foot Drop
Hindawi Publishing Corporation Case Reports in Orthopedics Volume 2016, Article ID 9024368, 3 pages http://dx.doi.org/10.1155/2016/9024368 Case Report An Unusual Presentation of Right-Sided Sciatica with Foot Drop Fergus J. McCabe1 and John P. McCabe2 1 School of Medicine, NUI Galway, Galway, Ireland 2Department of Trauma & Orthopaedics, Galway University Hospitals, Galway, Ireland Correspondence should be addressed to Fergus J. McCabe; [email protected] Received 25 January 2016; Revised 6 April 2016; Accepted 17 April 2016 Academic Editor: Koichi Sairyo Copyright © 2016 F. J. McCabe and J. P. McCabe. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Rarely, sciatica is of extraspinal aetiology. By compressing the sciatic nerve, swelling of the short external rotators of the hip can cause sciatica. Uncommon anatomical relationships between the sciatic nerve and local muscles may potentiate this compressive effect. In this report, we describe the presentation of right sciatica and foot drop resulting from both extreme local constriction and unusual anatomical variation of the right sciatic nerve. 1. Introduction stand, walk, sit, and in particular drive. There was no specific history of trauma to the area. Sitting aggravated the pain. The sciatic nerve is the major motor and sensory nerve of The pain and discomfort were sharp, continual, and severe, the lower limb [1]. In the vast majority of cases, the sciatic disturbing the man’s sleep pattern and hindering his quality nerve enters the lower limb via the greater sciatic foramen, of life.