Late Pleistocene Proboscideans from Yangjiawan Caves in Pingxiang Of
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

UCLA UCLA Electronic Theses and Dissertations
UCLA UCLA Electronic Theses and Dissertations Title Music and Language in the Strophic Singing of the Zhuang Minority in Southern China Permalink https://escholarship.org/uc/item/4t0423bz Author Widman, John Harvey Publication Date 2019 Peer reviewed|Thesis/dissertation eScholarship.org Powered by the California Digital Library University of California UNIVERSITY OF CALIFORNIA Los Angeles Music and Language in the Strophic Singing of the Zhuang Minority in Southern China A dissertation submitted in partial satisfaction of the requirements for the degree Doctor of Philosophy in Ethnomusicology by John Widman 2019 © Copyright by John Widman 2019 ABSTRACT OF THE DISSERTATION Music and Language in the Strophic Singing of the Zhuang Minority in Southern China by John Widman Doctor of Philosophy in Ethnomusicology University of California, Los Angeles, 2019 Professor Helen M. Rees, Chair The Zhuang minority in Guangxi, China, are commonly celebrated for their “sea of songs.” While the epithet of being “good at singing and dancing” is an overused description applied to China’s fifty-five minorities, Zhuang do gather in parks during weekends and holidays and sing strophic songs outside their homes and on mountain tops to each other in annual festivals. Although a given geographic area typically features only one of these tunes, in many areas of Guangxi, these songs are the primary form of music-making. This kind of strophic singing, known throughout China as mountain songs (shan’ge), has a variety of names spread among the dialects of the eighteen million Tai-Kadai speakers who make up the Zhuang. The Zhuang who live along the Youjiang river valley call their traditional singing fien. -

Anisotropic Patterns of Liver Cancer Prevalence in Guangxi in Southwest China: Is Local Climate a Contributing Factor?
DOI:http://dx.doi.org/10.7314/APJCP.2015.16.8.3579 Anisotropic Patterns of Liver Cancer Prevalence in Guangxi in Southwest China: Is Local Climate a Contributing Factor? RESEARCH ARTICLE Anisotropic Patterns of Liver Cancer Prevalence in Guangxi in Southwest China: Is Local Climate a Contributing Factor? Wei Deng1&, Long Long2&*, Xian-Yan Tang3, Tian-Ren Huang1, Ji-Lin Li1, Min- Hua Rong1, Ke-Zhi Li1, Hai-Zhou Liu1 Abstract Geographic information system (GIS) technology has useful applications for epidemiology, enabling the detection of spatial patterns of disease dispersion and locating geographic areas at increased risk. In this study, we applied GIS technology to characterize the spatial pattern of mortality due to liver cancer in the autonomous region of Guangxi Zhuang in southwest China. A database with liver cancer mortality data for 1971-1973, 1990-1992, and 2004-2005, including geographic locations and climate conditions, was constructed, and the appropriate associations were investigated. It was found that the regions with the highest mortality rates were central Guangxi with Guigang City at the center, and southwest Guangxi centered in Fusui County. Regions with the lowest mortality rates were eastern Guangxi with Pingnan County at the center, and northern Guangxi centered in Sanjiang and Rongshui counties. Regarding climate conditions, in the 1990s the mortality rate of liver cancer positively correlated with average temperature and average minimum temperature, and negatively correlated with average precipitation. In 2004 through 2005, mortality due to liver cancer positively correlated with the average minimum temperature. Regions of high mortality had lower average humidity and higher average barometric pressure than did regions of low mortality. -

Download Article (PDF)
Advances in Economics, Business and Management Research, volume 29 International Conference on Innovations in Economic Management and Social Science (IEMSS 2017) Suitability Assessment of Karst Rocky Desertification Control Patterns in Karst Counties of Guangxi Zhuang Autonomous Region Yan Yan1, 2, a, Baoqing Hu1, 2, Deguang Wang3 1Key laboratory of Environment Change and Resources Use in Beibu Gulf, (Guangxi Teachers Education University), Ministry of Education, Nanning, Guangxi, China; 2Guangxi Key Laboratory of Earth Surface Processes and Intelligent Simulation, Nanning, Guangxi, China; 3Guangxi University of Finance and Economics, Nanning, Guangxi, China; [email protected] Key words: Karst area of Guangxi; control patterns; fuzzy comprehensive assessment; regionalization Abstract. The State Council officially replied the “planning framework of comprehensive controls for karst rocky desertification regions” during 11th Five Year Plan. The counties in Guangxi appointed by the planning framework were taken as the study objects. First, the index system of suitability assessment was established and the fuzzy comprehensive assessment was conducted. The control patterns were regionalized by a bottom up method. Second, the karst rocky desertification control patterns under different geographic background were collected and summarized. The control patterns were generalized by a top down method. Finally, the assessment results of the two methods were compared and adjusted. Then, the karst counties of Guangxi were regionalized and the suitable control patterns for karst counties were determined. Introduction In Guangxi, some successful rocky desertification control patterns and techniques have been formed. But these methods are developed under different ecological, social and economic conditions [1-2]. These patterns have promotional value theoretically, but their application scopes still need to be discussed [3]. -

Cassava in China Inad• Era of Change
, '. -.:. " . Ie'"d;~~aVa in China lnan• I j Era of Change A CBN Case Study with Farmers and Processors ~-- " '. -.-,'" . ,; . ):.'~. - ...~. ¡.;; i:;f;~ ~ ';. ~:;':. __ ~~,.:';.: GuyHenry an~ Reinhardt Howeler )28103 U.' '1'/ "'.'..,· •.. :¡g.l ... !' . ~ .. W()R~mG,~6t:UMENT 1§:º~~U'U~T'O~OIln1ernotlonol CeMe:r fer TropIcal AgrICultura No. 155 Cassava Biotechnolgy Network Cassava in China InaD• Era of Change A CBN Case Study with Farmers and Processors GuyHenry and Reinhardt Howeler Cover Photos: Top: Cassava processing in Southern China í Bottom: Farmer participatory research in Southern China I I Al! photos: Cuy Henry (ClAn, July-August, 1994 I I¡ ¡ ¡, I Centro Internacional de Agricultura Tropical, CIAT ! Intemational Center for Tropical Agriculwre I Apartado Aéreo 6713 Cali, Colombia G:IAT Working Document No. 155 Press fun: 100 Printed in Colombia june 1996 ! Correa citation: Henry, G.; Howeler, R. 1996. Cassava in China in an era of change. A CBN case study with farmers and processors. 31 July to 20 August, 1994. - Cali Colombia: Centro Internacional de Agricultura Tropical, 1996. 68 p. - (Working Document; no. 1 ~5) I Cassava in China in An Era of Change A CBN Case Study with farmers and processors in Guangdong, Guangxi and Hainan Provinces of Southern China By: Guy Henry and Reínhardt Howeler luly 31 - August 20, 1994 Case Study Team Members: Dr. Guy Henry (Economist) International Center for Tropical Agriculture (ClAn, Cal i, Colombia Dr. Reinharot Howeler (Agronomis!) Intemational Center for Tropical Agricultur<! (ClAn, Bangkok, Thailand Mr. Huang Hong Cheng (Director), Mr. Fang Baiping, M •. Fu Guo Hui 01 the Upland Crops Researcll Institute (UCRIl in Guangzhou. -

China Construction Bank Corporation 2013 Annual Report Builds Modern Living
LIVING BUILDS BUILDS ANNUAL REPORT 2013 REPORT ANNUAL MODERN MODERN CHINA CONSTRUCTION BANK CORPORATION 2013 ANNUAL REPORT BUILDS MODERN LIVING China Construction Bank Corporation, established in October 1954 and headquartered in Beijing, is a leading large-scale joint stock commercial bank in Mainland China with world-renowned reputation. The Bank was listed on Hong Kong Stock Exchange in October 2005 (stock code: 939) and listed on the Shanghai Stock Exchange in September 2007 (stock code: 601939). At the end of 2013, the Bank’s market capitalisation reached US$187.8 billion, ranking 5th among listed banks in the world. With 14,650 branches and sub-branches in Mainland China, the Bank provides services to 3,065,400 corporate customers and 291 million personal customers, and maintains close cooperative relationships with a significant number of high-end customers and leading enterprises of strategic industries in the Chinese economy. The Bank maintains overseas branches in Hong Kong, Singapore, Frankfurt, Johannesburg, Tokyo, Osaka, Seoul, New York, Ho Chi Minh City, Sydney, Melbourne, Taipei and Luxembourg, and owns various subsidiaries, such as CCB Asia, CCB International, CCB London, CCB Russia, CCB Dubai, CCB Europe, CCB Principal Asset Management, CCB Financial Leasing, CCB Trust and CCB Life. The Bank upholds its “customer-centric, market-oriented” business concept, adheres to its development strategy of “integration, multifunction and intensiveness”, and strives to provide customers with premium and all-round modern financial services by accelerating innovation of products, channels and service modes. With a number of core business indicators leading the market, the Bank vigorously promotes the development of emerging businesses including electronic banking, private banking, credit cards, cash management, and pension, while maintaining its traditional businesses advantages in infrastructure and housing finance. -

A Sociolinguistic Survey of the Dejing Zhuang Dialect Area
DigitalResources Electronic Survey Report 2012-036 ® A Sociolinguistic Survey of the Dejing Zhuang Dialect Area Eric M. Jackson Emily H.S. Jackson Shuh Huey Lau A Sociolinguistic Survey of the Dejing Zhuang Dialect Area Eric M. Jackson, Emily H.S. Jackson, Shuh Huey Lau SIL International ® In cooperation with the Guangxi Minorities Language and Scripts Work Commission 2012 SIL Electronic Survey Report 2012-036, October 2012 Copyright © 2012 Eric M. Jackson, Emily H.S. Jackson, Shuh Huey Lau, and SIL International ® All rights reserved Contents Acknowledgements Abstract 1 Dejing Zhuang: An introduction 1.1 Piecing together the ethnolinguistic situation 1.2 Demographics and non-linguistic background 1.2.1 Geography 1.2.2 Ethnicity 1.2.3 Economy and commerce 1.2.4 Education 1.3 Linguistic background 1.3.1 The language development situation 1.3.2 Linguistic variation according to published accounts 1.4 Other background information 2 Purpose of the current field study 2.1 Research questions 2.2 Concepts, indicators, and instruments 3 Methods of the current field study 3.1 Selection of datapoints 3.2 Survey instruments 3.2.1 Wordlists 3.2.2 Recorded Text Tests 3.2.3 Post-RTT Individual Sociolinguistic Questionnaire 3.2.4 Group Sociolinguistic Questionnaire 3.2.5 Leader sociolinguistic questionnaire 3.3 Official partnership 3.4 Research ethics 4 Results and interpretation 4.1 Wordlist results: String edit distance analysis 4.2 Wordlist results: Tone systems 4.3 RTT results 4.3.1 RTT results: Factors in interpretation ii 4.3.2 RTT results: Interpretation -

Rebellion of the Cadres: the 1967 Implosion of the Chinese Party-State
Rebellion of the Cadres: The 1967 Implosion of the Chinese Party-State Andrew G. Walder* ABSTRACT Accounts of the tumultuous initial phase of the Cultural Revolution portray party-state cadres primarily as targets of a popular insurgency. Cadres in Party and government organs in fact were themselves in widespread rebellion against their superiors after October 1966, and rebel cadres were a major force in the national wave of power seizures that destroyed the civilian state in early 1967. The rebellion was a form of bureaucratic politics in a setting characterized by rapidly shifting signals and high uncertainty, in which the rebels’ motives were generated after the onset of the Cultural Revolution. Cadres played a central role in the destruction of the political institutions to which their vested interests were inextricably linked. he Cultural Revolution initially impressed scholars as a window on inequal- Tity and group conflict in a type of society long characterized as totalitarian. Observers immediately noted evidence that a series of social constituencies mo- bilized to advance their claims.1 Sent-down youth, contract and temporary work- ers, students from politically stigmatized households, demobilized soldiers, and other groups organized to press their interests and make demands against Party authorities.2 Mass insurgencies that seemed to reflect underlying group tensions * The author gratefully acknowledges the substantive and editorial suggestions of the editors, Anita Chan and Jonathan Unger, and the critical comments of two reviewers. National Science Foundation Grant SBS-1021134, “Political Movements in an Authoritarian Hierarchy,” supported some of the research reported in this article. 1. Michel Oksenberg, “Occupational Groups in Chinese Society and the Cultural Revolution,” and Ezra F. -

Conservation Recommendations for Oryza Rufipogon Griff. in China
www.nature.com/scientificreports OPEN Conservation recommendations for Oryza rufpogon Grif. in China based on genetic diversity analysis Junrui Wang1,6, Jinxia Shi2,6, Sha Liu1, Xiping Sun3, Juan Huang1,4, Weihua Qiao1,5, Yunlian Cheng1, Lifang Zhang1, Xiaoming Zheng1,5* & Qingwen Yang1,5* Over the past 30 years, human disturbance and habitat fragmentation have severely endangered the survival of common wild rice (Oryza rufpogon Grif.) in China. A better understanding of the genetic structure of O. rufpogon populations will therefore be useful for the development of conservation strategies. We examined the diversity and genetic structure of natural O. rufpogon populations at the national, provincial, and local levels using simple sequence repeat (SSR) markers. Twenty representative populations from sites across China showed high levels of genetic variability, and approximately 44% of the total genetic variation was among populations. At the local level, we studied fourteen populations in Guangxi Province and four populations in Jiangxi Province. Populations from similar ecosystems showed less genetic diferentiation, and local environmental conditions rather than geographic distance appeared to have infuenced gene fow during population genetic evolution. We identifed a triangular area, including northern Hainan, southern Guangdong, and southwestern Guangxi, as the genetic diversity center of O. rufpogon in China, and we proposed that this area should be given priority during the development of ex situ and in situ conservation strategies. Populations from less common ecosystem types should also be given priority for in situ conservation. Common wild rice (Oryza rufpogon Grif.) is the putative progenitor of Asian cultivated rice, one of the most important food crops in the world. -

World Bank Document
SFG2246 REV World Bank Financed -Guangxi Rural Poverty Alleviation Pilot Project Public Disclosure Authorized Public Disclosure Authorized Social Assessment Report Public Disclosure Authorized Guangxi Foreign Capital Poverty Reduction Project Management Center Public Disclosure Authorized June 2016 Table of Contents Executive Summary ......................................................................................................................................... 1 1 Introduction ....................................................................................................................................... 9 1.1 China’s National Poverty Alleviation Strategy ....................................................................................... 9 1.2 Guangxi’s Poverty Alleviation Strategy ................................................................................................ 10 1.3 Summary Project Description ................................................................................................................ 12 1.4 Policy Framework .................................................................................................................................... 13 1.4.1 Farmer Professional Cooperatives ............................................................................................... 13 1.4.2 Poverty Alleviation through Agricultural Industrialization .......................................................... 14 1.4.3 Land Transactions ......................................................................................................................... -

Agricultural Bank of China Corporate Social Responsibility Report 2018
Agricultural Bank of China Corporate Social Responsibility Report 2018 March 2019 Preface Time measures the progress of pioneers, and witnesses the dream of strivers. Time elapses in sweat and gains, and renews in yearning and hope. For China, 2018 is the 40th anniversary of the reform and opening up. For Agricultural Bank of China, 2018 is a year for deepening its reform and development, and the first year for comprehensively implementing the guiding principles of the 19th CPC National Congress. Standing at a new starting point, we held high the great banner of socialism with Chinese characteristics, firmly implemented Xi Jinping Thought on Socialism with Chinese Characteristics for a New Era, and spared no efforts to embark on a new journey of reform and development for ABC in the new era. Efforts never fail and dreams have never been closer. In 2018, we adhered to the philosophies of “giving priority to CSR, benefiting the general public, shouldering responsibilities, and promoting the well-being of society”. We seized the trend, led the future, run forward and never stopped. With wisdom and perseverance, we composed a beautiful song of “Sannong” service. With sweat and struggle, we drew a gorgeous painting of promoting the high-quality development of the real economy. With performance and responsibility, we vigorously answered to the call of “The Three” tough battles with efforts and pioneering spirit. We keep young in the elapsing time. In 2018, we held fast to the objective of “being a responsible bank”, further improved the social responsibility image, and continued to polish the social responsibility culture, to forge social responsibility into the beacon, the flag, the culture, and the source of motivation for ABC, and the soul leading and propping ABC’s reform and development. -

The Quest for Zhuang Identity: Cultural Politics of Promoting the Buluotuo Cultural Festival in Guangxi, China
THE QUEST FOR ZHUANG IDENTITY: CULTURAL POLITICS OF PROMOTING THE BULUOTUO CULTURAL FESTIVAL IN GUANGXI, CHINA SOMRAK CHAISINGKANANONT NATIONAL UNIVERSITY OF SINGAPORE 2014 THE QUEST FOR ZHUANG IDENTITY: CULTURAL POLITICS OF PROMOTING THE BULUOTUO CULTURAL FESTIVAL IN GUANGXI, CHINA SOMRAK CHAISINGKANANONT (B.A. HISTORY (2nd Class Hons.) THAMMASAT University, M.A. ANTHROPOLOGY, THAMMASAT University) A THESIS SUBMITTED FOR THE DEGREE OF DOCTOR OF PHILOSOPHY DEPARTMENT OF SOUTHEAST ASIAN STUDIES NATIONAL UNIVERSITY OF SINGAPORE 2014 i DECLARATION I hereby declare that the thesis is my original work and it has been written by me in its entirety. I have duly acknowledged all the sources of information which have been used in the thesis. This thesis has also not been submitted for any degree in any university previously. Somrak Chaisingkananont 22 August 2014 ii TABLE OF CONTENTS Declaration ii Acknowledgements v Summary vii List of Maps ix List of Figures x Notes on Transliteration and Abbreviations xii Chapter 1 Introduction 1 Chapter 2 Becoming “Zhuang zu”: Notion of Ethnicity as Cultural Politics 27 2.1 Guangxi as the Southern Borderland: A Narrative of Place and People 32 at the Empire‟s Margin 2.2 Zhonghua Minzu: The Rise of the New Chinese National Identity 40 2.2.1 Nation Building and Deployment of Ethnology 44 2.2.2 The Politics of Naming: Ethnonymic Polemics During Wartime 48 2.2.3 The Communist Party‟s Policy on Ethnic Minorities 51 2. 3 The Making of the Socialist “unified, multinational state” 55 2.3.1 In the Name of “Zhuang”: -
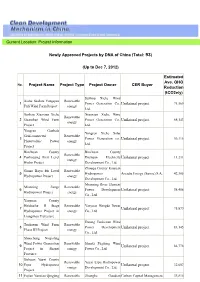
Current Location: Project Information Newly Approved Projects by DNA of China (Total: 93) (Up to Dec 7, 2012) Project Name Proje
Current Location: Project Information Newly Approved Projects by DNA of China (Total: 93) (Up to Dec 7, 2012) Estimated Ave. GHG No. Project Name Project Type Project Owner CER Buyer Reduction (tCO2e/y) Suzhou Xiehe Wind Xiehe Suzhou Yongqiao Renewable 1 Power Generation Co., Unilateral project 71,569 Fuli Wind Farm Project energy Ltd. Suzhou Xiaoxian Xiehe Xiaoxian Xiehe Wind Renewable 2 Guanshan Wind Farm Power Generation Co., Unilateral project 68,143 energy Project Ltd. Yongren Ganbala Yongren Xiehe Solar Grid-connected Renewable 3 Power Generation co., Unilateral project 56,116 Photovoltaic Power energy Ltd. Project Binchuan County Binchuan County Renewable 4 Pankoujing First Level Dachuan Electricity Unilateral project 11,231 energy Hydro Project Development Co., Ltd. Zhouqu County Kaiyuan Gansu Boyu 4th Level Renewable 5 Hydropower Arcadia Energy (Suisse) S.A. 42,305 Hydropower Project energy Development Co., Ltd. Mianning River Element Mianning Sange Renewable 6 Power Development Unilateral project 58,406 Hydropower Project energy Co., Ltd. Yanyuan County Heishuihe II Stage Renewable Yanyuan Hengda Power 7 Unilateral project 75,879 Hydropower Project in energy Co., Ltd. Liangshan Prefecture Datang Tuokexun Wind Tuokexun Wind Farm Renewable 8 Power Development Unilateral project 83,145 Phase III Project energy Co., Ltd. Shuocheng Niujialing Wind Power Generation Renewable Shanxi Fuguang Wind 9 Unilateral project 86,778 Project in Shanxi energy Power Co., Ltd. Province Sichuan Yuexi County Renewable Yuexi Erpu Hydropower 10 Erpu Hydropower Unilateral project 32,607 energy Development Co., Ltd. Project 11 Fujian Yunxiao Qingjing Renewable Zhonghe Guodian Carbon Capital Management, 33,810 Wind Farm Project energy Zhangzhou Energy Co., Inc.