Bioactive Potential of Minor Italian Olive Genotypes from Apulia, Sardinia and Abruzzo
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Report Di Gestione Dell'esercizio 2011
REPORT DI GESTIONE DELL’ESERCIZIO 2011 IV EDIZIONE A CURA DELL’UFFICIO INNOVAZIONE E QUALITA’ PRESENTAZIONE La quarta edizione del bilancio sociale dell’Università di Macerata - riferita all’anno 2011 – è il risultato di un lavoro corale dell’Ateneo. Lo dico con soddisfazione perché essere giunti alla quarta edizione ci dice che abbiamo sviluppato conoscenze e competenze preziose indispensabili ai fini del controllo, del reporting e soprattutto del miglioramento. Basta confrontare, anche superficialmente, questa edizione con le precedenti per rendersi conto di quanta strada è stata fatta, e in meglio. Il bilancio sociale integrale offre diverse opportunità. Anzitutto ci sollecita a considerare sempre il nostro lavoro come un tassello di un progetto collettivo, animato da valori e obiettivi comuni. Avere questa coscienza di sé significa avere maggiori possibilità di affrontare insieme le sfide e le indubbie criticità che contrassegnano il sistema universitario italiano, specie in questa fase di ripiegamento e di riorganizzazione. Il bilancio sociale ci interroga, ci presenta un’immagine, pur parziale, della nostra identità. Ci dice chi siamo e che cosa facciamo per continuare a fare con responsabilità e impegno il nostro lavoro. La nostra visione – l’umanesimo che innova – comunica una identità profonda, una focalizzazione scientifica e accademica peculiarissima in Italia. Il bilancio sociale 2011 ne dà ampia testimonianza, misurando, attraverso criteri e indicatori, i principali risultati. Questa nuova edizione rivela anche i passi che sono stati compiuti per migliorare l’utilizzo dello “strumento” bilancio sociale. La maggiore omogeneità ne è uno dei risultati più apprezzabili. Il bilancio è un utile strumento di comunicazione, mette in collegamento tutti i portatori di interesse all’interno dell’Ateneo e consente di instaurare con i soggetti “esterni” un confronto e un dialogo importanti ai fini dell’autovalutazione e delle scelte di politica accademica. -
![Sanna Alessandra-Clinica Neurologica [File.Pdf]](https://docslib.b-cdn.net/cover/2654/sanna-alessandra-clinica-neurologica-file-pdf-22654.webp)
Sanna Alessandra-Clinica Neurologica [File.Pdf]
F ORMATO EUROPEO PER IL CURRICULUM V I T A E INFORMAZIONI PERSONALI Nome SANNA ALESSANDRA Indirizzo VIA TEDDE 14 Telefono 3294061216 Fax E-mail [email protected] Nazionalità ITALIANA Data di nascita 01/12/1972 ESPERIENZA LAVORATIVA 1998 MEDICO PRESSO LA GUARDIA MEDICA TURISTICA DI BUDONI , NUORO (GIUGNO - AGOSTO ; INCARICO TRIMESTRALE ). 2000 Medico Volontario presso l’ambulatorio Caritas di Stubbla, Kosovo (due settimane). 2002 Medico presso la guardia medica di Ittiri, Sassari (aprile-giugno;incarico trimestrale). 2002 Medico presso la guardia medica turistica di Stintino, Sassari (luglio- settembre; incarico trimestrale). 2003 Medico presso la guardia medica di Ittiri, Sassari (dicembre-febbraio; incarico trimestrale). 2003 Medico presso la guardia medica turistica di Fertilia, Sassari (luglio- settembre; incarico trimestrale). Dal 2006 al 2008, assegnista di ricerca presso l’Università di Sassari; Dal Marzo 2003 all’Ottobre 2007, consulente neurologa presso l’A.I.A.S. Dal 1 luglio 2008 al 31/08/09 neurologa presso il poliambulatorio di Bonorva . Pagina 1 - Curriculum vitae di Per ulteriori informazioni: [ SANNA ALESSANDRA ] www.cedefop.eu.int/transparency www.europa.eu.int/comm/education/index_it.html www.eurescv-search.com Dal 23/08/04 al 31/08/09 dirigente medico branca psichiatria presso il CSM di Alghero (12 ore/settimana). Dal gennaio 2008 a tutt’oggi esperta presso il tribunale di sorveglianza di Sassari. Dal 23/04/09 al 07/09/09 dirigente medico neurologo presso l’AOU di Sassari Dal 08/09/09 a tutt’oggi, Dirigente Medico Neurologo presso l’Asl. 1 di Sassari con contratto a tempo indeterminato (dal Gennaio 2016 AOU di Sassari) Socia collaboratrice dell'Associazione “Nido delle Mamme” dal 2014 ISTRUZIONE E FORMAZIONE 1991: Diploma di Maturità Classica Luglio 1997: Laurea in medicina e chirurgia con votazione di 110/110 e lode. -

Tuscany & Umbria
ITALY Tuscany & Umbria A Guided Walking Adventure Table of Contents Daily Itinerary ........................................................................... 4 Tour Itinerary Overview .......................................................... 10 Tour Facts at a Glance ........................................................... 12 Traveling To and From Your Tour .......................................... 14 Information & Policies ............................................................ 17 Italy at a Glance ..................................................................... 19 Packing List ........................................................................... 24 800.464.9255 / countrywalkers.com 2 © 2015 Otago, LLC dba Country Walkers Travel Style This small-group Guided Walking Adventure offers an authentic travel experience, one that takes you away from the crowds and deep in to the fabric of local life. On it, you’ll enjoy 24/7 expert guides, premium accommodations, delicious meals, effortless transportation, and local wine or beer with dinner. Rest assured that every trip detail has been anticipated so you’re free to enjoy an adventure that exceeds your expectations. And, with our optional Flight + Tour ComboCombo, Florence PrePre----tourtour Extension and Rome PostPost----TourTour Extension to complement this destination, we take care of all the travel to simplify the journey. Refer to the attached itinerary for more details. Overview A walk in the sweeping hills of Tuscany and Umbria is a journey into Italy’s artistic and agricultural heart. Your path follows history, from Florence—where your tour commences—to Siena—an important art center distinguished by its remarkable cathedral—and on to Assisi to view the art treasures of the Basilica of St. Francis. Deep in Umbria, you view Gubbio’s stunning Palazzo dei Consolo and move on to the mosaics decorating Orvieto’s Gothic cathedral. Your stay in the Roman town of Spello—known for its medieval frescoes— inspires with aesthetic balance and timeless charm. -

How Best to Respond? Expert Meeting Djibuti, 8-10 November 2011
RefugeesRefugees andand asylumasylum--seekersseekers inin distressdistress atat seasea –– howhow bestbest toto respondrespond?? ExpertExpert meetingmeeting DjibutiDjibuti,, 88--1010 NovemberNovember 20112011 AUTHORITIES INVOLVED IN RESCUE AT SEA Central Directorate Strategic Coordination Navy Navy CoCoastast Guard Operational guidance in high seas Operational guidance for S.A.R. events Guardia di Finanza Police & Carabinieri Operational guidance in territorial waters Close shore line patrolling PrincipalPrincipal FlowsFlows towardstowards ItalyItaly From TUNISIA EGADI ISLANDS TUNISI TUNISIA LINOSA LAMPEDUSA SOUSSE MADIJA * Up to 5 November DATA ON LANDINGS YEARS LANDINGS MEN WOMEN MINOR TOTAL 2009 39 391 1 7 399 2010 51 560 2 52 614 2011* 512 26.682 235 1.102 28.019** **Landing in Lampedusa 25.714 Landing in Linosa 429 In 2011 have been arrested 73 smugglers and facilitators and 337 boats have been confiscated. In the 2010, were arrested only 7 persons and 19 boats were confiscated. Modus Operandi from Tunisia • By zodiac or wooden boat, of about 4 to 15 meters in length with 3 to 279 persons aboard (on a boat of 12 meters in length) • By fishing boats of 15/25 meters in length (maximum 344 persons aboard a boat of 15 meters in length) • Principally young males • Many trips are self-organized • Nocturnal departure • The cost is about 1,500/2,000 dinars • The Tunisians, generally, claim to want to reach northern Europe From LIBYA SICILY LINOSA LAMPEDUSA TRIPOLI ZUARA MISRATAH LIBYA * Up to 26 may DATA ON LANDINGS YEARS LANDINGS MEN WOMEN MINOR TOTAL 2009 55 4,928 896 466 6,290 2010 9 279 10 57 346 2011* 99 23.137 3.016 1.985 28.318 In 2011 have been arrested 51 smugglers and facilitators and have been confiscated 60 boats. -

“Please You Cannot Ignore Any Longer the Fact That These Animals Have
NAME AFFILIATION Diana Reiss, Ph.D. New York Aquarium and Columbia University “Please you Lori Marino, Ph.D. Emory University Prof. PatholoGy, School of Medicine, Sam RidGway, D.V.M., Ph.D. cannot ignore University of California Director of the New York Aquarium Paul J. Boyle, Ph.D. any longer the and The Ocean Project fact that these Randall Wells, Ph.D. Chicago ZooloGical Society animals have Hal Whitehead, Ph.D. Dalhousie University, Canada Louis M. Herman, Ph.D. University of Hawaii very large Paul NachtiGall, Ph.D. University of Hawaii brains, highly Richard Connor. Ph.D. University of Massachusetts, Dartmouth Adjunct Professor, Scripps Institution of William F. Perrin, Ph.D. developed OceanoGraphy Executive Vice Provost, Florida International societies, social DouGlas Wartzok, Ph.D. University relationships and Robin W. Baird, Ph.D. Cascadia Research Collective sophisticated Christopher W. Clark, Ph.D. Cornell University cognitive Adam Pack, Ph.D. University of Hawaii William E. Evans, Ph.D. University of Notre Dame abilities.” James Estes, Ph.D. Adjunct Professor, UC Santa Cruz – Richard Connor, Ph.D., University of Massachusetts Laureline Meynier, Ph.D. Massey University, New Zealand David Bain University of WashinGton Center for Marine Biodiversity and Sarah Mesnick, Ph.D. Conservation Janet Mann, Ph.D. GeorGetown University Leszek Karczmarski, Ph.D. Texas A&M University Corrie Allen, Ph.D. Lone Wolf Bioscience Giovanni Di Guardo. DVM, University of Teramo, Italy Dipl. ECVP Dr. L.P. Verstraten University of Amsterdam Cornelis Hazevoet Ph.D. Lisbon National Museum of Natural History Giuseppe Notarbartolo di Tethys Research Institute, Italy Sciara Ph.D. Dr. Victoria Turner Appin Scientific Limited Heidi Lyn, Ph.D. -

Ph.D. Thesis Morrone
UNIVERSITA’ DEGLI STUDI DI PARMA Department of Food Science Ph. D. in Food Science and Technology Cycle XXVII Relationship between environmental features and extra virgin olive oil in north Sardinia Ph. D. Coordinator: Chiar.mo Prof. Furio Brighenti Tutor: Chiar.mo Prof. Andrea Fabbri Co-Tutors: Dott.ssa Annalisa Rotondi Dott. Tommaso Ganino Ph.D. Student: Lucia Morrone Lucia Morrone, 2015 Relationship between environmental features and extra virgin olive oil in north Sardinia PhD Thesis in Food Science and Technology. XXVII Cycle, University of Parma, ITALY. Thesis Supervisors: Prof. Andrea Fabbri – Department of Food Science, University of Parma Dr. Annalisa Rotondi – Institute of BIoMETeorology of the National Research Council (IBIMET – CNR), Bologna Dr. Tommaso Ganino - Department of Food Science, University of Parma PhD Coordinator : Prof. Furio Brighenti – Department of Food Science, University of Parma II To Maurizio III IV Preface and Acknowledgements The agri-food sector is a strategic asset for Italy, representing the 8,7% of GDP. The significance of this sector is not merely economic, even if the agri-food sector is an important item of GDP and it has always a positive mark in export. As a matter of fact, the agri-food sector has both a social and an environmental impact. In this regard, the valorisation of Italian agri food productions, the so- called Made-in-Italy Agri-Food, assumes a crucial importance. The extra virgin olive oil is one of the products that most personify the image of the Made-in-Italy Agri-Food. Notwithstanding a lot of people think at the Italian virgin olive oil like a one and definite product, it is a product having hundreds of chemical and sensory shades. -

Multiple Sclerosis in the Campania Region (South Italy): Algorithm Validation and 2015–2017 Prevalence
International Journal of Environmental Research and Public Health Article Multiple Sclerosis in the Campania Region (South Italy): Algorithm Validation and 2015–2017 Prevalence Marcello Moccia 1,* , Vincenzo Brescia Morra 1, Roberta Lanzillo 1, Ilaria Loperto 2 , Roberta Giordana 3, Maria Grazia Fumo 4, Martina Petruzzo 1, Nicola Capasso 1, Maria Triassi 2, Maria Pia Sormani 5 and Raffaele Palladino 2,6 1 Multiple Sclerosis Clinical Care and Research Centre, Department of Neuroscience, Reproductive Science and Odontostomatology, Federico II University, Via Sergio Pansini 5, 80131 Naples, Italy; [email protected] (V.B.M.); [email protected] (R.L.); [email protected] (M.P.); [email protected] (N.C.) 2 Department of Public Health, Federico II University, 80131 Naples, Italy; [email protected] (I.L.); [email protected] (M.T.); raff[email protected] (R.P.) 3 Campania Region Healthcare System Commissioner Office, 80131 Naples, Italy; [email protected] 4 Regional Healthcare Society (So.Re.Sa), 80131 Naples, Italy; [email protected] 5 Biostatistics Unit, Department of Health Sciences, University of Genoa, 16121 Genoa, Italy; [email protected] 6 Department of Primary Care and Public Health, Imperial College, London SW7 2AZ, UK * Correspondence: [email protected] or [email protected]; Tel./Fax: +39-081-7462670 Received: 21 April 2020; Accepted: 12 May 2020; Published: 13 May 2020 Abstract: We aim to validate a case-finding algorithm to detect individuals with multiple sclerosis (MS) using routinely collected healthcare data, and to assess the prevalence of MS in the Campania Region (South Italy). To identify individuals with MS living in the Campania Region, we employed an algorithm using different routinely collected healthcare administrative databases (hospital discharges, drug prescriptions, outpatient consultations with payment exemptions), from 1 January 2015 to 31 December 2017. -
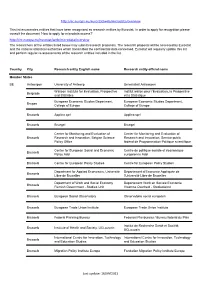
Eurostat: Recognized Research Entity
http://ec.europa.eu/eurostat/web/microdata/overview This list enumerates entities that have been recognised as research entities by Eurostat. In order to apply for recognition please consult the document 'How to apply for microdata access?' http://ec.europa.eu/eurostat/web/microdata/overview The researchers of the entities listed below may submit research proposals. The research proposal will be assessed by Eurostat and the national statistical authorities which transmitted the confidential data concerned. Eurostat will regularly update this list and perform regular re-assessments of the research entities included in the list. Country City Research entity English name Research entity official name Member States BE Antwerpen University of Antwerp Universiteit Antwerpen Walloon Institute for Evaluation, Prospective Institut wallon pour l'Evaluation, la Prospective Belgrade and Statistics et la Statistique European Economic Studies Department, European Economic Studies Department, Bruges College of Europe College of Europe Brussels Applica sprl Applica sprl Brussels Bruegel Bruegel Center for Monitoring and Evaluation of Center for Monitoring and Evaluation of Brussels Research and Innovation, Belgian Science Research and Innovation, Service public Policy Office fédéral de Programmation Politique scientifique Centre for European Social and Economic Centre de politique sociale et économique Brussels Policy Asbl européenne Asbl Brussels Centre for European Policy Studies Centre for European Policy Studies Department for Applied Economics, -

Economic Survey of North Sardinia 2014
Economic Survey of North Sardinia 2014 Economic Survey of North Sardinia 2014 PAGE 1 Economic Survey of North Sardinia 2014 Introduction Through the publication of the Economic Survey of North Sardinia, the Chamber of Commerce of Sassari aims to provide each year an updated and detailed study concerning social and economic aspects in the provinces of Sassari and Olbia-Tempio and, more generally, in Sardinia . This survey is addressed to all the entrepreneurs and institutions interested in the local economy and in the potential opportunities offered by national and foreign trade. Indeed, this analysis takes into account the essential aspects of the entrepreneurial activity (business dynamics, agriculture and agro-industry, foreign trade, credit, national accounts, manufacturing and services, etc…). Moreover, the analysis of the local economy, compared with regional and national trends, allows to reflect on the future prospects of the territory and to set up development projects. This 16th edition of the Survey is further enriched with comments and a glossary, intended to be a “guide” to the statistical information. In data-processing, sources from the Chamber Internal System – especially the Business Register – have been integrated with data provided by public institutions and trade associations. The Chamber of Commerce wishes to thank them all for their collaboration. In the last years, the Chamber of North Sardinia has been editing and spreading a version of the Survey in English, in order to reach all the main world trade operators. International operators willing to invest in this area are thus supported by this Chamber through this deep analysis of the local economy. -

ROGER II of SICILY a Ruler Between East and West
. ROGER II OF SICILY A ruler between east and west . HUBERT HOUBEN Translated by Graham A. Loud and Diane Milburn published by the press syndicate of the university of cambridge The Pitt Building, Trumpington Street, Cambridge cb2 1rp, United Kingdom cambridge university press The Edinburgh Building, Cambridge, cb2 2ru,UK 40 West 20th Street, New York, ny 10011-4211, USA 477 Williamstown Road, Port Melbourne, vic 3207, Australia Ruiz de Alarcon´ 13, 28014 Madrid, Spain Dock House, The Waterfront, Cape Town 8001, South Africa http://www.cambridge.org Originally published in German as Roger II. von Sizilien by Wissenschaftliche Buchgesellschaft, Darmstadt, 1997 and C Wissenschaftliche Buchgesellschaft, Darmstadt, 1997 First published in English by Cambridge University Press 2002 as Roger II of Sicily English translation C Cambridge University Press 2002 This book is in copyright. Subject to statutory exception and to the provisions of relevant collective licensing agreements, no reproduction of any part may take place without the written permission of Cambridge University Press. Printed in the United Kingdom at the University Press, Cambridge Typeface Bembo 10/11.5 pt. System LATEX 2ε [TB] A catalogue record for this book is available from the British Library Library of Congress Cataloguing in Publication data Houben, Hubert. [Roger II. von Sizilien. English] Roger II of Sicily: a ruler between east and west / Hubert Houben; translated by Graham A. Loud and Diane Milburn. p. cm. Translation of: Roger II. von Sizilien. Includes bibliographical references and index. isbn 0 521 65208 1 (hardback) isbn 0 521 65573 0 (paperback) 1. Roger II, King of Sicily, d. -

UNIVERSITY of TERAMO (Italy) * * * * *
“International Co-operation Against Trans-national Financial Organized Crime” Master – University of Teramo UNIVERSITY OF TERAMO (Italy) * * * * * CALL FOR APPLICATIONS MASTER IN “I NTERNATIONAL CO-OPERATION AGAINST TRANS -NATIONAL FINANCIAL ORGANIZED CRIME ” Academic Year 2009-2010 DURATION 2. environmental crimes, such as unlawful 1 year divided in three parts: building, pollution, trafficking in waste, Part 1 : starting from Spring 2010 at the University of radioactive slag, animals’ and plants’ Teramo, consisting in 200 hours of didactic activity. endangered species The didactic activity will be held with the following 3. corruption and bribery schedule: Thursday afternoon, Friday morning and 4. brand and copyright counterfeiting afternoon and Saturday morning; this calendar might 5. pharmaceutical counterfeiting and food undergo variations. adulteration 6. money laundering and financial crime Part 2 : starting after the end of part 1 an internship 7. terrorism and terrorism financing with international or national relevant institutions, or 8. terrorism and biotechnologies a research programme with scientific institutions. 9. cyber crime Admission to the internship will be granted to those and the substantive action undertaken by the who have attended at least 80% of the didactic activity following national and international organisations: hours and passed the intermediary written - DNA ( Direzione Nazionale Antimafia ) examinations. - EUROJUST ( European Union’s Judicial The internship period is not compulsory for Cooperation Unit ) – Permanent Mission of Italy candidates. - EUROPOL Part 3 : Candidates that have successfully concluded - INTERPOL their internship period must elaborate and defend a - OCSE ( Organisation for Economic Co-operation final thesis (not less than 100 pages). and Development ) - Permanent Mission of Italy Candidates not attending an internship must elaborate - OLAF ( European Anti-Fraud Office ) and discuss a final thesis (not less than 300 pages). -

The North-South Divide in Italy: Reality Or Perception?
CORE Metadata, citation and similar papers at core.ac.uk EUROPEAN SPATIAL RESEARCH AND POLICY Volume 25 2018 Number 1 http://dx.doi.org/10.18778/1231-1952.25.1.03 Dario MUSOLINO∗ THE NORTH-SOUTH DIVIDE IN ITALY: REALITY OR PERCEPTION? Abstract. Although the literature about the objective socio-economic characteristics of the Italian North- South divide is wide and exhaustive, the question of how it is perceived is much less investigated and studied. Moreover, the consistency between the reality and the perception of the North-South divide is completely unexplored. The paper presents and discusses some relevant analyses on this issue, using the findings of a research study on the stated locational preferences of entrepreneurs in Italy. Its ultimate aim, therefore, is to suggest a new approach to the analysis of the macro-regional development gaps. What emerges from these analyses is that the perception of the North-South divide is not consistent with its objective economic characteristics. One of these inconsistencies concerns the width of the ‘per- ception gap’, which is bigger than the ‘reality gap’. Another inconsistency concerns how entrepreneurs perceive in their mental maps regions and provinces in Northern and Southern Italy. The impression is that Italian entrepreneurs have a stereotyped, much too negative, image of Southern Italy, almost a ‘wall in the head’, as also can be observed in the German case (with respect to the East-West divide). Keywords: North-South divide, stated locational preferences, perception, image. 1. INTRODUCTION The North-South divide1 is probably the most known and most persistent charac- teristic of the Italian economic geography.