Potential Impacts of Projected Sea-Level Rise on Sea Turtle Rookeries
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Between Australia and New Guinea-Ecological and Cultural
Geographical Review of Japan Vol. 59 (Ser. B), No. 2, 69-82, 1986 Between Australia and New Guinea-Ecological and Cultural Diversity in the Torres Strait with Special Reference to the Use of Marine Resources- George OHSHIMA* The region between lowland Papua and the northern tip of the Australian continent presents a fascinating panorama of ecological, cultural and socio-economic diversity. In lowland Papua and on its associated small islands such as Saibai, Boigu and Parama, a combination of coastal forests and muddy shores dominates the scene, whereas the Torres Strait Islands of volcanic and limestone origin, together with raised coral islands and their associated reef systems present a range of island ecosystems scattered over a broad territory some 800km in extent. Coralline habitats extend south wards to the Cape York Peninsula and some parts of Arnhem Land. Coupled with those ecological diversities within a relatively small compass across the Torres Strait, the region has evoked important questions concerning the archaeological and historical dichotomy between Australian hunter-gatherers and Melanesian horticulturalists. This notion is also reflectd in terms of its complex linguistic, ethnic and political composition. In summarizing the present day cultural diversity of the region, at least three major components emerge: hunter-gatherers in the Australian Northern Territory, Australian islanders and tribal Papuans. Historically, these groups have interacted in complex ways and this has resulted in an intricate intermingling of cultures and societies. Such acculturation processes operating over thousands of years make it difficult to isolate meaning ful trends in terms of "core-periphery" components of the individual cultures. -

Net Primary Productivity, Global Climate Change and Biodiversity In
This file is part of the following reference: Fuentes, Mariana Menezes Prata Bezerra (2010) Vulnerability of sea turtles to climate change: a case study within the northern Great Barrier Reef green turtle population. PhD thesis, James Cook University. Access to this file is available from: http://eprints.jcu.edu.au/11730 Vulnerability of sea turtles to climate change: A case study with the northern Great Barrier Reef green turtle population PhD thesis submitted by Mariana Menezes Prata Bezerra FUENTES (BSc Hons) February 2010 For the degree of Doctor of Philosophy School of Earth and Environmental Sciences James Cook University Townsville, Queensland 4811 Australia “The process of learning is often more important than what is being learned” ii Statement of access I, the undersigned, author of this work, understand that James Cook University will make this thesis available for use within the University Library, via the Australian Theses Network, or by other means allow access to users in other approved libraries. I understand that as an unpublished work, a thesis has significant protection under the Copyright Act and beyond this, I do not wish to place any restriction access to this thesis. _______________________ ____________________ Signature, Mariana Fuentes Date iii Statement of sources declaration I declare that this thesis is my own work and has not been submitted in any form for another degree or diploma at any university or other institution of tertiary education. Information derived from the published or unpublished work of others has been duly acknowledged in the text and a list of references is given. ________________________ _____________________ Signature, Mariana Fuentes Date iv Statement of contribution of others Research funding: . -

Land & Sea Management Strategy
LAND & SEA MANAGEMENT STRATEGY for TORRES STRAIT Funded under the Natural Heritage Trust LAND & SEA MANAGEMENT STRATEGY for TORRES STRAIT Torres Strait NRM Reference Group with the assistance of Miya Isherwood, Michelle Pollock, Kate Eden and Steve Jackson November 2005 An initiative funded under the Natural Heritage Trust program of the Australian Government with support from: the Queensland Department of Natural Resources & Mines the Australian Government Department of the Environment and Heritage and the Torres Strait Regional Authority. Prepared by: Bessen Consulting Services FOREWORD The Torres Strait region is of enormous significance from a land and sea management perspective. It is a geographically and ecologically unique region, which is home to a culturally distinct society, comprised of over 20 different communities, extending from the south-western coast of Papua New Guinea (PNG) to the northern tip of Cape York Peninsula. The Torres Strait is also a very politically complex area, with an international border and a Treaty governance regime that poses unique challenges for coordinating land and sea management initiatives. Torres Strait Islander and Aboriginal people, both within and outside of the region, have strong and abiding connections with their land and sea country. While formal recognition of this relationship has been achieved through native title and other legal processes, peoples’ aspirations to sustainably manage their land and sea country have been more difficult to realise. The implementation of this Strategy, with funding from the Australian Government’s Natural Heritage Trust initiative, presents us with an opportunity to progress community-based management in our unique region, through identifying the important land and sea assets, issues, information, and potential mechanisms for supporting Torres Strait communities to sustainably manage their natural resources. -

New Life-Span Records of the Brown Booby Sula Leucogaster
Hennicke et al.: Brown Booby life-span 125 NEW LIFE-SPAN RECORDS OF THE BROWN BOOBY SULA LEUCOGASTER JANOS C. HENNICKE1,2, BRIAN KING3†, DAVID DRYNAN4, LISA J. HARDY4, ANTHONY STOKES5 & SASCHA TAYLOR3 1Current address: Centre d’Études Biologiques de Chizé, CEBC – CNRS, Villiers-en-Bois, France 2Department of Ecology and Conservation, University of Hamburg, Germany ([email protected]) 3Queensland Parks and Wildlife Service (Marine Parks), Queensland Dept. of Environment and Resource Management, Brisbane, Australia †deceased 30 March 1992 4Australian Bird & Bat Banding Scheme, Dept. of Sustainability, Environment, Water, Population and Communities, Canberra, Australia 5P.O. Box 1553, Townsville, Queensland, Australia Received 15 August 2012, accepted 18 September 2012 Seabirds are exposed to a highly variable marine environment in (3) On May 10, 2009, a dead bird with a leg ring was found in which prey availability is unpredictable, patchy and ephemeral the Brown Booby colony at the “Dales”, Christmas Island, Indian (Ashmole 1971, Hunt & Schneider 1987, Shealer 2002). Ocean (10.5°S, 105.6°E). The bird was already decomposed to Consequently, seabirds have evolved a number of distinct life bones and feathers, but the carcass was complete, and the bones history traits to cope with those conditions, such as low fecundity, were still largely held together by ligaments. It had been banded deferred maturity and high adult survival (e.g. Furness & Monaghan at approximately the same location on August 16, 1985. Time and 1987, Weimerskirch 2002). An important life history trait of cause of death are unknown but, given the condition of the carcass, seabirds is their longevity. -

Recovery Plan for the Bramble Cay Melomys Melomys Rubicola Prepared by Peter Latch
Recovery Plan for the Bramble Cay Melomys Melomys rubicola Prepared by Peter Latch Title: Recovery Plan for the Bramble Cay Melomys Melomys rubicola Prepared by: Peter Latch © The State of Queensland, Environmental Protection Agency, 2008 Copyright protects this publication. Except for purposes permitted by the Copyright Act, reproduction by whatever means is prohibited without the prior written knowledge of the Environmental Protection Agency. Inquiries should be addressed to PO Box 15155, CITY EAST QLD 4002. Copies may be obtained from the: Executive Director Conservation Services Environmental Protection Agency PO Box 15155 CITY EAST Qld 4002 Disclaimer: The Australian Government, in partnership with the Environmental Protection Agency facilitates the publication of recovery plans to detail the actions needed for the conservation of threatened native wildlife. The attainment of objectives and the provision of funds may be subject to budgetary and other constraints affecting the parties involved, and may also be constrained by the need to address other conservation priorities. Approved recovery actions may be subject to modification due to changes in knowledge and changes in conservation status. Publication reference: Latch, P. 2008.Recovery Plan for the Bramble Cay Melomys Melomys rubicola. Report to Department of the Environment, Water, Heritage and the Arts, Canberra. Environmental Protection Agency, Brisbane. 2 Contents Page No. Executive Summary 4 1. General information 5 Conservation status 5 International obligations 5 Affected interests 5 Consultation with Indigenous people 5 Benefits to other species or communities 5 Social and economic impacts 5 2. Biological information 5 Species description 5 Life history and ecology 7 Description of habitat 7 Distribution and habitat critical to the survival of the species 9 3. -

Autonomy Rights in Torres Strait: from Whom, for Whom, for Or Over What?
Autonomy rights in Torres Strait: From whom, for whom, for or over what? W.G. Sanders and W.S. Arthur No. 215/2001 ISSN 1036-1774 ISBN 0 7315 2650 3 Will Sanders is a Fellow and Bill Arthur a Research Fellow at the Centre for Aboriginal Economic Policy Research, The Australian National University. CENTRE FOR ABORIGINAL ECONOMIC POLICY RESEARCH DISCUSSION PAPER NO. 215 iii Table of contents Abbreviations and acronyms ...............................................................................iv Summary .............................................................................................................v Acknowledgments ................................................................................................v Introduction ........................................................................................................ 1 Calls for autonomy, and existing and proposed governance structures ................ 4 From whom? ....................................................................................................... 8 For whom? .......................................................................................................... 9 For or over what? .............................................................................................. 12 Autonomy, Australian federalism, and regional ethnic diversity: A concluding comment ...................................................................................... 15 Notes................................................................................................................ -

Determination 2016 (No 2)
Australian Capital Territory Public Place Names (Throsby) Determination 2016 (No 2) Disallowable instrument DI2016-211 made under the Public Place Names Act 1989, s 3 (Minister to determine names) 1 Name of instrument This instrument is the Public Place Names (Throsby) Determination 2016 (No 2). 2 Commencement This instrument commences on the day after notification. 3 Determination of Place Names I determine the names listed in the schedule as the names of the corresponding public places indicated in the associated plan in the schedule. Dorte Ekelund Delegate of the Minister 24 August 2016 Authorised by the ACT Parliamentary Counsel—also accessible at www.legislation.act.gov.au SCHEDULE See section 3 of the Public Place Names (Throsby) Determination 2016 (No 2) Division of Throsby – Native Fauna The location of the public places with the following names is indicated on the associated plan. NAME ORIGIN SIGNIFICANCE Kaigas Street Kaigas On Mabuiag Island in the Torres Strait the word ‘Kaigas’ has the Traditional languages meaning of ‘Shovelnose shark’ in the of the Torres Strait English language. Islands. Kunapalari Kunapalari Frog Amphibian; frog Street Neobatrachus The Kunapalari Frog is a common kunapalari species in the southwest and central zones of Western Australia. The frog (Wheatbelt Frog) is usually found on clay soils. It is a moderate-sized robust species with relatively short limbs and protruding eyes. The Kunapalari Frog breeds after autumn or winter rain. It mates in milky pools where the resulting tadpoles are usually hidden from view. The call is a distinctive long, low trill. Melomys Circuit Bramble Cay melomys Mammal, native rodent Melomys rubicola A small, nocturnal native rodent unique to Bramble Cay, the most (Mosaic-tailed rat) northern island of the Great Barrier Reef in the northern Torres Strait. -

Foraging Ecology of Masked Boobies: the Importance of Intrinsic and Extrinsic Factors
Foraging ecology of Masked Boobies: The importance of intrinsic and extrinsic factors by Julia Sommerfeld Diploma in Zoology, University of Hamburg Submitted in fulfilment of the requirements for the degree of Doctor of Philosophy Institute for Marine and Antarctic Studies, University of Tasmania Research and Technology Centre, University of Kiel June 2013 i ZUSAMMENFASSUNG Um ein fundiertes Verständnis über die Ernährungsökologie mariner Top-Prädatoren zu erlangen, ist es erforderlich, sowohl die Effekte intrinsischer Eigenschaften des Tieres zu untersuchen, als auch extrinsche, umweltbedingte Faktoren präzise zu bestimmen. In der vorliegenden Arbeit wurden dazu die Bewegungs- und Aktivitätsmuster kükenaufziehender Maskentölpel Sula dactylatra über zwei Brutzeiten auf Phillip Island (Norfolk Island Gruppe), Australien (Südpazifik) mit Hilfe von GPS- und Tauch- Beschleunigungs-Datenloggern aufgezeichnet. Ziel dieser Arbeit ist es Folgendes zu bestimmen: (I) Beeinflussen intrinsche Faktoren, wie zum Beispiel unterschiedliche Körpergewichte und Größe innerhalb desselben Geschlechts, die Bewegungsmuster, Flug- und Tauchverhalten von Männchen und Weibchen? (II) Verändern Adulttiere ihre Bewegungsmuster und ihr Tauchverhalten im Laufe der Untersuchungszeit und falls ja, steht dies im Verhältnis zu den Änderungen in der Nahrungsverteilung? (III) Die Bestimmung der Genaugigkeit der First-Passage Time (FPT) Analyse – eine Methode die zunehmend verwendet wird, um spezifische Nahrungssuchgebiete (Area-restricted-search (ARS) Zonen) anhand von GPS-Daten zu identifizieren. (I) Maskentölpel weisen einen umgekehrten Geschlechtsdimorphismus auf (d.h. Weibchen sind größer als Männchen). Die Ergebnisse der vorliegenden Studie belegen, dass der Wechsel zwischen kurzen und langen Beutezügen bei Weibchen abhängig vom Körpergewicht ist und wahrscheinlich dazu dient, abwechselnd Nahrung für das Küken zu erbeuten (kurze Beutezüge) bzw. zur Selbsternährung dient (lange Beutezüge). -

Nesting Green Turtles of Torres Strait
Final Report Nesting green turtles of Torres Strait Mark Hamann, Justin Smith, Shane Preston and Mariana Fuentes Nesting green turtles of Torres Strait Final report Mark Hamann1, Justin Smith1, Shane Preston1 and Mariana Fuentes1 1 College of Marine and Environmental Sciences, James Cook University Supported by the Australian Government’s National Environmental Research Program Project 2.1 Marine turtles and dugongs of the Torres Strait © JCU National Library of Australia Cataloguing-in-Publication entry: 978-1-925088-62-5 This report should be cited as: Hamann, M., Smith, J. Preston S. and Fuentes, M.M.P.B. (2015) Nesting green turtles of Torres Strait. Report to the National Environmental Research Program. Reef and Rainforest Research Centre Limited, Cairns (15pp.). Published by the Reef and Rainforest Research Centre on behalf of the Australian Government’s National Environmental Research Program (NERP) Tropical Ecosystems (TE) Hub. The Tropical Ecosystems Hub is part of the Australian Government’s Commonwealth National Environmental Research Program. The NERP TE Hub is administered in North Queensland by the Reef and Rainforest Research Centre Limited (RRRC). The NERP Tropical Ecosystem Hub addresses issues of concern for the management, conservation and sustainable use of the World Heritage listed Great Barrier Reef (GBR) and its catchments, tropical rainforests including the Wet Tropics World Heritage Area (WTWHA), and the terrestrial and marine assets underpinning resilient communities in the Torres Strait, through the generation and transfer of world-class research and shared knowledge. This publication is copyright. The Copyright Act 1968 permits fair dealing for study, research, information or educational purposes subject to inclusion of a sufficient acknowledgement of the source. -

Great Barrier Reef
PAPUA 145°E 150°E GULF OF PAPUA Dyke NEW GUINEA O Ackland W Bay GREAT BARRIER REEF 200 E Daru N S General Reference Map T Talbot Islands Anchor Cay A Collingwood Lagoon Reef N Bay Saibai Port Moresby L Reefs E G Island Y N E Portlock Reefs R A Torres Murray Islands Warrior 10°S Moa Boot Reef 10°S Badu Island Island 200 Eastern Fields (Refer Legend below) Ashmore Reef Strait 2000 Thursday 200 Island 10°40’55"S 145°00’04"E WORLD HERITAGE AREA AND REGION BOUNDARY ait Newcastle Bay Endeavour Str GREAT BARRIER REEF WORLD HERITAGE AREA Bamaga (Extends from the low water mark of the mainland and includes all islands, internal waters of Queensland and Seas and Submerged Lands Orford Bay Act exclusions.) Total area approximately 348 000 sq km FAR NORTHERN Raine Island MANAGEMENT AREA GREAT BARRIER REEF REGION CAPE Great Detached (Extends from the low water mark of the mainland but excludes lburne Bay he Reef Queensland-owned islands, internal waters of Queensland and Seas S and Submerged Lands Act exclusions.) Total area approximately 346 000 sq km ple Bay em Wenlock T GREAT BARRIER REEF MARINE PARK (Excludes Queensland-owned islands, internal waters of Queensland River G and Seas and Submerged Lands Act exclusions.) Lockhart 4000 Total area approximately 344 400 sq km Weipa Lloyd Bay River R GREAT BARRIER REEF MARINE PARK 12°59’55"S MANAGEMENT AREA E 145°00’04"E CORAL SEA YORK GREAT BARRIER REEF PROVINCE Aurukun River A (As defined by W.G.H. -

Year in Review
YEAR IN REVIEW Saving our Reef and its marine life for future generations The Great Barrier Reef Foundation extends its deepest ‘Reef Wonderland’ artwork by Melanie Hava, Mamu Aboriginal respect and recognition to all Traditional Owners of the woman, Dugulbarra and Great Barrier Reef and its Catchments, as First Nations Waribarra family groups, from Peoples holding the hopes, dreams, traditions and the Johnstone River catchment of the Wet Tropics of Far North cultures of the Reef. Queensland and the adjoining More than 70 Traditional Owner groups have deep and Great Barrier Reef sea country. enduring connections spanning the length of the Reef along the Queensland coastline and beyond, from the Torres Strait Islands in the north to Bundaberg in the south. Cover image credit: Gary Cranitch, Queensland Museum. CONTENTS Our Mission 2 From the Managing Director and Chair 4 Our Impact 6 Australia’s Largest Ever Reef Partnership 8 Saving Endangered Species 10 Innovation In All We Do 16 Restoring Critical Reef Habitats 22 Thriving Reefs, Thriving Communities 26 Building Global Reef Resilience 30 Our Supporters 32 Board and Committees 37 2019-20 YEAR IN REVIEW 1 OUR MISSION We bring together the best minds and technologies to help solve the challenges facing the Great Barrier Reef. And we challenge everyone — from the boardroom to the beach — to stand up, unite, and take an active part in its conservation. The Great Barrier Reef is an This unique icon is the largest We should be proud that it’s our irreplaceable ecosystem, home to living thing on Earth, with rich responsibility to look after it. -
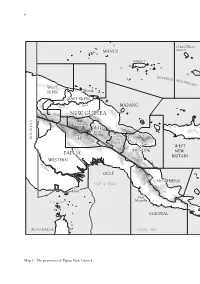
I2I Text Paste Up
vi ST MATTHIAS MANUS GROUP MANUS I BIS MARCK ARCH IPEL AGO WEST SEPIK Wewak EAST SSEPIKEPIK River Sepik MADANG NEW GUINEA ENGA W.H. Mt Hagen M Goroka a INDONESIA S.H. rk ha E.H. m R Lae WEST MOROBEMOR PAPUA NEW BRITAIN WESTERN F ly Ri ver GULF NORTHERNOR N Gulf of Papua Daru Port Torres Strait Moresby CENTRAL AUSTRALIA CORAL SEA Map 1: The provinces of Papua New Guinea vii 0 300 kilometres 0 150 miles NEW IRELAND PACIFIC OCEAN NEW IRELAND Rabaul BOUGAINVILLE I EAST Arawa NEW BRITAIN Panguna SOLOMON SEA SOLOMON ISLANDS D ’EN N TR E C A S T E A U X MILNE BAY I S LOUISIADE ARCHIPELAGO © Carto ANU 05-031 viii W ALLAC E'S LINE SUNDALAND WALLACEA SAHULLAND 0 500 km © Carto ANU 05-031b Map 2: The prehistoric continent of Sahul consisted of the continent of Australia and the islands of New Guinea and Tasmania. ix 141 o E 142°E 143°E 144°E 144°15'E PAPUA NEW GUINEA Parama I. 9°S Daru I. 9°S Bramble Cay Missionary Reef Boigu I. Anchor Cay s f Dauan I. e East Cay Saibai I. e Deliverance I. R Stephens I. r Darnley I. Kerr (Kiss) Islet o ri ar W Yorke I. Turu Cay Mabuiag I. Yam I. Murray Islands 10°S Coconut I. 10°S Badu I. Moa I. TORRES Kubin St Pauls Warraber (Sue) I. STRAIT OUTER ISLANDS V INNER Hammond I. Thursday I. CORAL SEA ISLANDS Horn I. N Prince of Authority area Wales I.