Food Web Ecology: Models and Application to Conservation
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Untangling Taxonomy: a DNA Barcode Reference Library for Canadian Spiders
Molecular Ecology Resources (2016) 16, 325–341 doi: 10.1111/1755-0998.12444 Untangling taxonomy: a DNA barcode reference library for Canadian spiders GERGIN A. BLAGOEV, JEREMY R. DEWAARD, SUJEEVAN RATNASINGHAM, STEPHANIE L. DEWAARD, LIUQIONG LU, JAMES ROBERTSON, ANGELA C. TELFER and PAUL D. N. HEBERT Biodiversity Institute of Ontario, University of Guelph, Guelph, ON, Canada Abstract Approximately 1460 species of spiders have been reported from Canada, 3% of the global fauna. This study provides a DNA barcode reference library for 1018 of these species based upon the analysis of more than 30 000 specimens. The sequence results show a clear barcode gap in most cases with a mean intraspecific divergence of 0.78% vs. a min- imum nearest-neighbour (NN) distance averaging 7.85%. The sequences were assigned to 1359 Barcode index num- bers (BINs) with 1344 of these BINs composed of specimens belonging to a single currently recognized species. There was a perfect correspondence between BIN membership and a known species in 795 cases, while another 197 species were assigned to two or more BINs (556 in total). A few other species (26) were involved in BIN merges or in a combination of merges and splits. There was only a weak relationship between the number of specimens analysed for a species and its BIN count. However, three species were clear outliers with their specimens being placed in 11– 22 BINs. Although all BIN splits need further study to clarify the taxonomic status of the entities involved, DNA bar- codes discriminated 98% of the 1018 species. The present survey conservatively revealed 16 species new to science, 52 species new to Canada and major range extensions for 426 species. -

Dipterists Digest
Dipterists Digest 2019 Vol. 26 No. 1 Cover illustration: Eliozeta pellucens (Fallén, 1820), male (Tachinidae) . PORTUGAL: Póvoa Dão, Silgueiros, Viseu, N 40º 32' 59.81" / W 7º 56' 39.00", 10 June 2011, leg. Jorge Almeida (photo by Chris Raper). The first British record of this species is reported in the article by Ivan Perry (pp. 61-62). Dipterists Digest Vol. 26 No. 1 Second Series 2019 th Published 28 June 2019 Published by ISSN 0953-7260 Dipterists Digest Editor Peter J. Chandler, 606B Berryfield Lane, Melksham, Wilts SN12 6EL (E-mail: [email protected]) Editorial Panel Graham Rotheray Keith Snow Alan Stubbs Derek Whiteley Phil Withers Dipterists Digest is the journal of the Dipterists Forum . It is intended for amateur, semi- professional and professional field dipterists with interests in British and European flies. All notes and papers submitted to Dipterists Digest are refereed. Articles and notes for publication should be sent to the Editor at the above address, and should be submitted with a current postal and/or e-mail address, which the author agrees will be published with their paper. Articles must not have been accepted for publication elsewhere and should be written in clear and concise English. Contributions should be supplied either as E-mail attachments or on CD in Word or compatible formats. The scope of Dipterists Digest is: - the behaviour, ecology and natural history of flies; - new and improved techniques (e.g. collecting, rearing etc.); - the conservation of flies; - reports from the Diptera Recording Schemes, including maps; - records and assessments of rare or scarce species and those new to regions, countries etc.; - local faunal accounts and field meeting results, especially if accompanied by ecological or natural history interpretation; - descriptions of species new to science; - notes on identification and deletions or amendments to standard key works and checklists. -

Aranei: Linyphiidae)
Arthropoda Selecta 29(4): 495–498 © ARTHROPODA SELECTA, 2020 A new species of the genus Silometopus Simon, 1926 from the Caspian Lowland, Russia (Aranei: Linyphiidae) Íîâûé âèä Silometopus Simon, 1926 èç Ïðèêàñïèéñêîé íèçìåííîñòè (Ðîññèÿ) (Aranei: Linyphiidae) Andrei V. Tanasevitch, Tatyana V. Grushko1 À.Â. Òàíàñåâè÷, Ò.Â. Ãðóøêî A.N. Severtsov Institute of Ecology and Evolution, Russian Academy of Sciences, Leninsky prospekt 33, Moscow 119071, Russia. E-mail: [email protected]; [email protected] Институт проблем экологии и эволюции им. А.Н. Северцова РАН, Ленинский проспект 33, Москва 119071, Россия. KEY WORDS: taxonomy, spiders, Arachnida, Erigoninae, Russian Plain, arid fauna. КЛЮЧЕВЫЕ СЛОВА: таксономия, пауки, Arachnida, Erigoninae, Русская равнина, аридная фауна. ABSTRACT. A new erigonine species, Silometo- Ponomarev & Dvadnenko [2012], Ponomarev et al. pus elton sp.n., is described from a semi-desert in the [2018]; Tanasevitch [1987, 1993], Tanasevitch & northwestern part of the Caspian Lowland. The spe- Piterkina [2007], etc. cies somewhat resembles the trans-Palaearctic S. re- The genus Silometopus Simon, 1926 presently in- ussi (Thorell, 1871), but can easily be distinguished cludes 17 species, most of which are restricted to Eu- by the structure of the distal suprategular apophysis rope [World Spider Catalog, 2020]. One species, S. and the shape of the palpal tibia. Based on its habitat incurvatus (O. Pickard-Cambridge, 1873), has been preferences, Silometopus elton sp.n. can preliminary recorded from the Caspian Lowland [Piterkina, 2009], be characterized as an element of a semi-desert intra- and one more, S. crassipedis Tanasevitch et Piterkina, zonal fauna of the southeastern part of the Russian 2007, has been described from that territory [Ta- Plain. -

Taxonomic Notes on Agroeca (Araneae, Liocranidae)
Arachnol. Mitt. 37: 27-30 Nürnberg, Juli 2009 Taxonomic notes on Agroeca (Araneae, Liocranidae) Torbjörn Kronestedt Abstract: Agroeca gaunitzi Tullgren, 1952 is stated here to be a junior synonym of A. proxima (O. P.-Cambridge, 1871). The illustrations of the male palp attributed to A. proxima in papers by Tullgren of 1946 and 1952 in fact show A. inopina O. P.-Cambridge, 1886. The record of A. inopina from Finland, quite outside its known distribution range, was based on a misidentification. It is argued that the type species of the genus Agroeca Westring, 1861 should be A. proxima (O. P.-Cambridge, 1871), not A. brunnea (Blackwall, 1833) as currently applied. Protagroeca Lohmander, 1944 is placed as an objective synonym of Agroeca Westring, 1861. Key words: Agroeca gaunitzi, synonyms, type species On the identity of Agroeca gaunitzi Tullgren, 1952 Because Agroeca gaunitzi still appears as a valid Agroeca gaunitzi was described based on a single nominal species (HELSDINGEN 2009, PLATNICK male from the southern part of Swedish Lapland 2009), a re-study of the holotype was undertaken in (TULLGREN 1952). No additional specimens have order to clarify its identity. As a result, the following since been assigned to this nominal species and it conclusions were reached: was not mentioned in the most recent taxonomical revision of the genus (GRIMM 1986) nor in the 1. The holotype of Agroeca gaunitzi is a male of latest treatment of the family in Sweden (ALM- Agroeca proxima, thus making the former a junior QUIST 2006). synonym, syn. n. According to the original description, A. gau- 2. -

Spiders (Araneae) of Churchill, Manitoba: DNA Barcodes And
Blagoev et al. BMC Ecology 2013, 13:44 http://www.biomedcentral.com/1472-6785/13/44 RESEARCH ARTICLE Open Access Spiders (Araneae) of Churchill, Manitoba: DNA barcodes and morphology reveal high species diversity and new Canadian records Gergin A Blagoev1*, Nadya I Nikolova1, Crystal N Sobel1, Paul DN Hebert1,2 and Sarah J Adamowicz1,2 Abstract Background: Arctic ecosystems, especially those near transition zones, are expected to be strongly impacted by climate change. Because it is positioned on the ecotone between tundra and boreal forest, the Churchill area is a strategic locality for the analysis of shifts in faunal composition. This fact has motivated the effort to develop a comprehensive biodiversity inventory for the Churchill region by coupling DNA barcoding with morphological studies. The present study represents one element of this effort; it focuses on analysis of the spider fauna at Churchill. Results: 198 species were detected among 2704 spiders analyzed, tripling the count for the Churchill region. Estimates of overall diversity suggest that another 10–20 species await detection. Most species displayed little intraspecific sequence variation (maximum <1%) in the barcode region of the cytochrome c oxidase subunit I (COI) gene, but four species showed considerably higher values (maximum = 4.1-6.2%), suggesting cryptic species. All recognized species possessed a distinct haplotype array at COI with nearest-neighbour interspecific distances averaging 8.57%. Three species new to Canada were detected: Robertus lyrifer (Theridiidae), Baryphyma trifrons (Linyphiidae), and Satilatlas monticola (Linyphiidae). The first two species may represent human-mediated introductions linked to the port in Churchill, but the other species represents a range extension from the USA. -

Sciomyzidae Et Phaeomyiidae De La Manche. Premier
Sciomyzidae et Phaeomyiidae de la Manche Premier catalogue Parmi les 132 familles de diptères dénombrées en Europe, celle des Sciomyzidae n’évoque sans doute que peu de choses aux fidèles lecteurs de L’Argiope. Dans notre faune, aucun de ses représentants n’est véritablement remarquable, ni même connu sous un nom populaire. Non floricoles, ces insectes à faible mobilité sont pour le moins discrets, et seul un observateur attentif et patient saura alors les déceler dans leur milieu de reproduction, notamment du fait d’un comportement singulier : les sciomyzes Sepedon sphegea se tiennent habituellement dans la végétation basse la tête orientée vers le bas ! Dans le petit monde des diptéristes français, les sciomyzes ont en revanche été rendus populaires par une importante publication. Dernier numéro en date consacré à une famille de diptères dans la série Faune de France, est paru en 1989 les « Diptères Sciomyzidae Euro-méditerranéens ». Plus de cinquante ans après SÉGUY (1934), Jean-Claude VALA présente dans cet ouvrage un état des connaissances détaillé quant à la biologie générale de ces insectes et pour de nombreuses espèces. Plusieurs espèces de sciomyzes ont été notamment étudiées de près en raison de leur importance dans la lutte contre certaines maladies humaines (distomatoses). En effet, certains parasites trématodes connus chez l’homme ont un cycle de développement qui nécessite un hôte intermédiaire spécifique parmi les mollusques aquatiques. Parasitoïdes ou prédateurs des mollusques aquatiques et terrestres, les sciomyzes s’opposent alors efficacement au développement de ces parasites. Paradoxalement, alors que la publication de VALA , en français, rend l’étude des sciomyzes beaucoup plus abordable que pour d’autres familles de mouches, ce travail n’a pas semblé entraîner un engouement particulier de la part des entomologistes amateurs puisque peu d’études ou de catalogues ont vu le jour depuis cette parution. -

Two New Cave-Dwelling Spider Species from the Moroccan High Atlas (Araneae: Liocranidae, Theridiidae)
Arachnologische Mitteilungen / Arachnology Letters 60: 12-18 Karlsruhe, September 2020 Two new cave-dwelling spider species from the Moroccan High Atlas (Araneae: Liocranidae, Theridiidae) Sylvain Lecigne, Josiane Lips, Soumia Moutaouakil & Pierre Oger doi: 10.30963/aramit6002 Abstract. During a scientific internship in a mountainous area near Agadir, several caves were prospected; the sampled material includ- ed several spider species. Morphological and taxonomical analysis revealed two new species to science: an anophthalmic troglobiont species, Agraecina agadirensis spec. nov. (description based on three females) and a new member of the genus Steatoda in the Mediter- ranean region, Steatoda ifricola spec. nov. (description based on both sexes). Diagnoses, drawings and photos are presented. Keywords: Agadir, comb-footed spiders, Ida Outanane Massif, spiny-legged sac spiders, taxonomy, troglobiont species Zusammenfassung. Zwei neue höhlenbewohnenden Spinnenarten aus dem marokkanischem Hohen Atlas (Araneae: Liocrani- dae, Theridiidae). Während eines wissenschaftlichen Praktikums im Gebirge nah Agadir wurden mehrere Höhlen besucht. Das dort gesammelte Material enthielt mehrere Spinnenarten. Die morphologische und taxonomische Untersuchung erbrachte zwei neue Arten für die Wissenschaft: eine augenlose troglobionte Art, Agraecina agadirensis spec. nov. (drei Weibchen) und eine neue Vertreterin der Gattung in der mediterranen Region, Steatoda ifricola spec. nov. (beide Geschlechter). Diagnosen, Zeichnungen und Fotos werden prä- sentiert. The araneofauna of Morocco is still only very partially known; triangulosa (Walckenaer, 1802), and in North Africa (S. vena- this is all the more so with respect to cave spiders. The present tor (Audouin, 1826)) (Nentwig et al. 2020, World Spider Cat- work illustrates this and provides the description of two new alog 2020). The genus Steatoda constitutes a group of spiders species for science. -
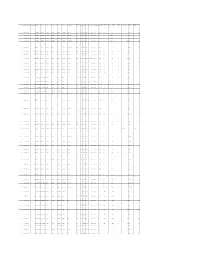
Appendix D: Summary of Accepted ECOTOX Papers
Effect Dur Dur Unit Dur Dur Unit Conc Conc Units Conc Value1 Purity CAS Number Chemical Name Species Number Phylum Class Order Family Genus Species Common Name Group Effect Meas Endpt1 Endpt2 Habitat Plant/Animal Media Orig Orig Preferred Preferred Type Conc Value1 Orig Orig Adjusted 1 298044 Disulfoton 4510 Chordata Mammalia Rodentia Muridae Rattus norvegicus Norway rat BEH AVO STIM NOAEL terrestrial Animal NONE 30 d 30 d F 2 mg/kg/d 1.96 2 298044 Disulfoton 351 Chordata Actinopterygii Perciformes Anabantidae Anabas testudineus Climbing perch BCM BCM LIPD LOAEL aquatic Animal FW 1 h 4.17E-02 d A 4 mg/L 4 3 298044 Disulfoton 351 Chordata Actinopterygii Perciformes Anabantidae Anabas testudineus Climbing perch BCM BCM PRCO LOAEL aquatic Animal FW 1 h 4.17E-02 d A 4 mg/L 4 4 298044 Disulfoton 351 Chordata Actinopterygii Perciformes Anabantidae Anabas testudineus Climbing perch BCM BCM LIPD LOAEL aquatic Animal FW 1 h 4.17E-02 d A 10.5 mg/L 10.5 5 298044 Disulfoton 4913 Chordata Mammalia Rodentia Muridae Mus musculus House mouse BCM BCM HXBT LOAEL terrestrial Animal NONE 3 d 3 d F 35.1 uM/kg 35.1 6 298044 Disulfoton 4510 Chordata Mammalia Rodentia Muridae Rattus norvegicus Norway rat BCM BCM GBCM LOAEL terrestrial Animal NONE 24 h 1 d F 0.26 mg/kg 0.234 7 298044 Disulfoton 4510 Chordata Mammalia Rodentia Muridae Rattus norvegicus Norway rat BCM BCM GBCM IC50 terrestrial Animal NONE 1 wk 7 d F 0.6 ppm 0.6 8 298044 Disulfoton 4510 Chordata Mammalia Rodentia Muridae Rattus norvegicus Norway rat BCM BCM GBCM NOAEL LOAEL terrestrial Animal NONE -

Nomenclatural Studies Toward a World List of Diptera Genus-Group Names
Nomenclatural studies toward a world list of Diptera genus-group names. Part V Pierre-Justin-Marie Macquart Evenhuis, Neal L.; Pape, Thomas; Pont, Adrian C. DOI: 10.11646/zootaxa.4172.1.1 Publication date: 2016 Document version Publisher's PDF, also known as Version of record Document license: CC BY Citation for published version (APA): Evenhuis, N. L., Pape, T., & Pont, A. C. (2016). Nomenclatural studies toward a world list of Diptera genus- group names. Part V: Pierre-Justin-Marie Macquart. Magnolia Press. Zootaxa Vol. 4172 No. 1 https://doi.org/10.11646/zootaxa.4172.1.1 Download date: 02. Oct. 2021 Zootaxa 4172 (1): 001–211 ISSN 1175-5326 (print edition) http://www.mapress.com/j/zt/ Monograph ZOOTAXA Copyright © 2016 Magnolia Press ISSN 1175-5334 (online edition) http://doi.org/10.11646/zootaxa.4172.1.1 http://zoobank.org/urn:lsid:zoobank.org:pub:22128906-32FA-4A80-85D6-10F114E81A7B ZOOTAXA 4172 Nomenclatural Studies Toward a World List of Diptera Genus-Group Names. Part V: Pierre-Justin-Marie Macquart NEAL L. EVENHUIS1, THOMAS PAPE2 & ADRIAN C. PONT3 1 J. Linsley Gressitt Center for Entomological Research, Bishop Museum, 1525 Bernice Street, Honolulu, Hawaii 96817-2704, USA. E-mail: [email protected] 2 Natural History Museum of Denmark, Universitetsparken 15, 2100 Copenhagen, Denmark. E-mail: [email protected] 3Oxford University Museum of Natural History, Parks Road, Oxford OX1 3PW, UK. E-mail: [email protected] Magnolia Press Auckland, New Zealand Accepted by D. Whitmore: 15 Aug. 2016; published: 30 Sept. 2016 Licensed under a Creative Commons Attribution License http://creativecommons.org/licenses/by/3.0 NEAL L. -

196 Arachnology (2019)18 (3), 196–212 a Revised Checklist of the Spiders of Great Britain Methods and Ireland Selection Criteria and Lists
196 Arachnology (2019)18 (3), 196–212 A revised checklist of the spiders of Great Britain Methods and Ireland Selection criteria and lists Alastair Lavery The checklist has two main sections; List A contains all Burach, Carnbo, species proved or suspected to be established and List B Kinross, KY13 0NX species recorded only in specific circumstances. email: [email protected] The criterion for inclusion in list A is evidence that self- sustaining populations of the species are established within Great Britain and Ireland. This is taken to include records Abstract from the same site over a number of years or from a number A revised checklist of spider species found in Great Britain and of sites. Species not recorded after 1919, one hundred years Ireland is presented together with their national distributions, before the publication of this list, are not included, though national and international conservation statuses and syn- this has not been applied strictly for Irish species because of onymies. The list allows users to access the sources most often substantially lower recording levels. used in studying spiders on the archipelago. The list does not differentiate between species naturally Keywords: Araneae • Europe occurring and those that have established with human assis- tance; in practice this can be very difficult to determine. Introduction List A: species established in natural or semi-natural A checklist can have multiple purposes. Its primary pur- habitats pose is to provide an up-to-date list of the species found in the geographical area and, as in this case, to major divisions The main species list, List A1, includes all species found within that area. -

ARTHROPODA Subphylum Hexapoda Protura, Springtails, Diplura, and Insects
NINE Phylum ARTHROPODA SUBPHYLUM HEXAPODA Protura, springtails, Diplura, and insects ROD P. MACFARLANE, PETER A. MADDISON, IAN G. ANDREW, JOCELYN A. BERRY, PETER M. JOHNS, ROBERT J. B. HOARE, MARIE-CLAUDE LARIVIÈRE, PENELOPE GREENSLADE, ROSA C. HENDERSON, COURTenaY N. SMITHERS, RicarDO L. PALMA, JOHN B. WARD, ROBERT L. C. PILGRIM, DaVID R. TOWNS, IAN McLELLAN, DAVID A. J. TEULON, TERRY R. HITCHINGS, VICTOR F. EASTOP, NICHOLAS A. MARTIN, MURRAY J. FLETCHER, MARLON A. W. STUFKENS, PAMELA J. DALE, Daniel BURCKHARDT, THOMAS R. BUCKLEY, STEVEN A. TREWICK defining feature of the Hexapoda, as the name suggests, is six legs. Also, the body comprises a head, thorax, and abdomen. The number A of abdominal segments varies, however; there are only six in the Collembola (springtails), 9–12 in the Protura, and 10 in the Diplura, whereas in all other hexapods there are strictly 11. Insects are now regarded as comprising only those hexapods with 11 abdominal segments. Whereas crustaceans are the dominant group of arthropods in the sea, hexapods prevail on land, in numbers and biomass. Altogether, the Hexapoda constitutes the most diverse group of animals – the estimated number of described species worldwide is just over 900,000, with the beetles (order Coleoptera) comprising more than a third of these. Today, the Hexapoda is considered to contain four classes – the Insecta, and the Protura, Collembola, and Diplura. The latter three classes were formerly allied with the insect orders Archaeognatha (jumping bristletails) and Thysanura (silverfish) as the insect subclass Apterygota (‘wingless’). The Apterygota is now regarded as an artificial assemblage (Bitsch & Bitsch 2000). -

20140620 Thesis Vanklink
University of Groningen Of dwarves and giants van Klink, Roel IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite from it. Please check the document version below. Document Version Publisher's PDF, also known as Version of record Publication date: 2014 Link to publication in University of Groningen/UMCG research database Citation for published version (APA): van Klink, R. (2014). Of dwarves and giants: How large herbivores shape arthropod communities on salt marshes. s.n. Copyright Other than for strictly personal use, it is not permitted to download or to forward/distribute the text or part of it without the consent of the author(s) and/or copyright holder(s), unless the work is under an open content license (like Creative Commons). The publication may also be distributed here under the terms of Article 25fa of the Dutch Copyright Act, indicated by the “Taverne” license. More information can be found on the University of Groningen website: https://www.rug.nl/library/open-access/self-archiving-pure/taverne- amendment. Take-down policy If you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediately and investigate your claim. Downloaded from the University of Groningen/UMCG research database (Pure): http://www.rug.nl/research/portal. For technical reasons the number of authors shown on this cover page is limited to 10 maximum. Download date: 29-09-2021 Of Dwarves and Giants How large herbivores shape arthropod communities on salt marshes Roel van Klink This PhD-project was carried out at the Community and Conservation Ecology group, which is part of the Centre for Ecological and Environmental Studies of the University of Groningen, The Netherlands.