Chapter 3 Natural Terrestrial Ecosystems
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
North America Other Continents
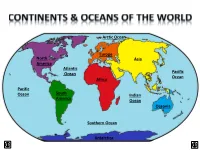
Arctic Ocean Europe North Asia America Atlantic Ocean Pacific Ocean Africa Pacific Ocean South Indian America Ocean Oceania Southern Ocean Antarctica LAND & WATER • The surface of the Earth is covered by approximately 71% water and 29% land. • It contains 7 continents and 5 oceans. Land Water EARTH’S HEMISPHERES • The planet Earth can be divided into four different sections or hemispheres. The Equator is an imaginary horizontal line (latitude) that divides the earth into the Northern and Southern hemispheres, while the Prime Meridian is the imaginary vertical line (longitude) that divides the earth into the Eastern and Western hemispheres. • North America, Earth’s 3rd largest continent, includes 23 countries. It contains Bermuda, Canada, Mexico, the United States of America, all Caribbean and Central America countries, as well as Greenland, which is the world’s largest island. North West East LOCATION South • The continent of North America is located in both the Northern and Western hemispheres. It is surrounded by the Arctic Ocean in the north, by the Atlantic Ocean in the east, and by the Pacific Ocean in the west. • It measures 24,256,000 sq. km and takes up a little more than 16% of the land on Earth. North America 16% Other Continents 84% • North America has an approximate population of almost 529 million people, which is about 8% of the World’s total population. 92% 8% North America Other Continents • The Atlantic Ocean is the second largest of Earth’s Oceans. It covers about 15% of the Earth’s total surface area and approximately 21% of its water surface area. -

Geography Notes.Pdf
THE GLOBE What is a globe? a small model of the Earth Parts of a globe: equator - the line on the globe halfway between the North Pole and the South Pole poles - the northern-most and southern-most points on the Earth 1. North Pole 2. South Pole hemispheres - half of the earth, divided by the equator (North & South) and the prime meridian (East and West) 1. Northern Hemisphere 2. Southern Hemisphere 3. Eastern Hemisphere 4. Western Hemisphere continents - the largest land areas on Earth 1. North America 2. South America 3. Europe 4. Asia 5. Africa 6. Australia 7. Antarctica oceans - the largest water areas on Earth 1. Atlantic Ocean 2. Pacific Ocean 3. Indian Ocean 4. Arctic Ocean 5. Antarctic Ocean WORLD MAP ** NOTE: Our textbooks call the “Southern Ocean” the “Antarctic Ocean” ** North America The three major countries of North America are: 1. Canada 2. United States 3. Mexico Where Do We Live? We live in the Western & Northern Hemispheres. We live on the continent of North America. The other 2 large countries on this continent are Canada and Mexico. The name of our country is the United States. There are 50 states in it, but when it first became a country, there were only 13 states. The name of our state is New York. Its capital city is Albany. GEOGRAPHY STUDY GUIDE You will need to know: VOCABULARY: equator globe hemisphere continent ocean compass WORLD MAP - be able to label 7 continents and 5 oceans 3 Large Countries of North America 1. United States 2. Canada 3. -

Seventh Grade Social Studies, Eastern Hemisphere Course Essential Questions (From Phase I Report)
Davison Community Schools ADVISORY CURRICULUM COUNCIL Phase II, April 2012 Seventh Grade Social Studies, Eastern Hemisphere Course Essential Questions (from Phase I report): 1. How does the past link to the present and determine the future? 2. How does the United States fit into the global world and how is the United States impacted by other nations? 3. What is the relationship between people and the Earth? 4. How can information be used to make informed decisions about understanding the world around us? Phase II Curriculum Unit: Geography Essential Questions: Essential Understanding: How do we use maps and other geographic tools Physical features impact where and how people to acquire information about the world? live What methods of inquiry and analysis do we use Geography impacts population patterns to answer geographic questions? Latitude and elevations impact climate zones How do we use themes, connections and spatial Geographers use 5 themes of geography to patterns to describe and study the Earth? describe regions or places on earth How do we compare and describe natural regions? Human and physical characteristics are used to How do we use human characteristics to study study the earth places on the Earth? Map skills are necessary for understanding information for a variety of maps Recognize distortion in a variety of map scale/projections Economic indicators (GDP, Literacy rate …..) are used to evaluate a country Globalization has created a global interdependence among people There are different purposes of world organizations Curriculum Standards G1.1.1 Explain and use a variety of maps, globes, and web based geography technology to study the world, including global, interregional, regional, and local scales. -

Smithsonian Miscellaneous Collections
-&? SMITHSONIAN MISCELLANEOUS COLLECTIONS VOLUME 82. NUMBER 6 THE PAST CLIMATE OF THE NORTH POLAR REGION BY EDWARD W. BERRY The Johns Hopkins University (Publication 3061) CITY OF WASHINGTON PUBLISHED BY THE SMITHSONIAN INSTITUTION APRIL 9, 1930 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOLUME 82, NUMBER 6 THE PAST CLIMATE OF THE NORTH POLAR REGION BY EDWARD W. BERRY The Johns Hopkins University Publication 306i i CITY OF WASHINGTON PUBLISHED BY THE SMITHSONIAN INSTITUTION APRIL 9, 1930 ZU £or& (gafttmore (prees BALTIMORE, MD., U. S. A. THE PAST CLIMATE OF THE NORTH POLAR REGION 1 By EDWARD W. BERRY THE JOHNS HOPKINS UNIVERSITY The plants, coal beds, hairy mammoth and woolly rhinoceros ; the corals, ammonites and the host of other marine organisms, chiefly invertebrate but including ichthyosaurs and other saurians, that have been discovered beneath the snow and ice of boreal lands have always made a most powerful appeal to the imagination of explorers and geologists. We forget entirely the modern whales, reindeer, musk ox, polar bear, and abundant Arctic marine life, and remember only the seemingly great contrast between the present and this subjective past. Nowhere on the earth is there such an apparent contrast between the present and geologic climates as in the polar regions and the mental pictures which have been aroused and the theories by means of which it has been sought to explain the fancied conditions of the past are all, at least in large part, highly imaginary. Occasionally a student like Nathorst (1911) has refused to be carried away by his imagination and has called to mind the mar- velously rich life of the present day Arctic seas, but for the most part those who have speculated on former climates have entirely ignored the results of Arctic oceanography. -

The World's Continents and Oceans the Seven Continents
7/29/2018 Identifying the Continents & Oceans of the World | Study.com Refer a friend Plans Courses Credit Degrees Schools Identifying the Continents & Oceans of the World In this lesson, we will learn about the location of the seven continents and the ve oceans of the world. We will identify their locations and highlight some of their Support key characteristics. The World's Continents and Oceans Geography is important. Imagine trying to describe Europe to someone who had no idea where in the world it is located. Geographical location provides us with context. It is important to know where the seven continents are located and what their characteristics are. It is also important to know where the world's ve oceans are. That is what we will do in this lesson. To begin with, look at the below map to familiarize yourself with the world's continents. The Seven Continents Geographers have divided the earth into seven continents or landmasses. Each of them is distinct, with its own distinct culture and characteristics. We'll begin in the Western Hemisphere of the earth, which includes North America and South America. North America On a map, North America is the northern of the two landmasses. For many of This map shows the seven continents in the world. us who are Americans, North America is easily recognizable. The Florida peninsula and the narrowing of land in the south are perhaps its key geographical features. Today, the countries of Canada, the U.S., Mexico, and the smaller Central American countries can be seen on a political map of North America. -

Ecosystem Diversity Report George Washington National Forest Draft EIS April 2011
Appendix E Ecosystem Diversity Report George Washington National Forest Draft EIS April 2011 U.S. Department of Agriculture Forest Service Southern Region Ecosystem Diversity Report George Washington National Forest April 2011 Appendix E Ecosystem Diversity Report George Washington National Forest Draft EIS April 2011 Table of Contents TABLE OF CONTENTS .............................................................................................................. I 1.0 INTRODUCTION.................................................................................................................. 1 2.0 ECOLOGICAL SUSTAINABILITY EVALUATION PROCESS ................................... 3 3.0 ECOLOGICAL SYSTEMS .................................................................................................. 7 3.1 Background and Distribution of Ecosystems ..................................................................................................... 7 North-Central Appalachian Acidic Swamp ............................................................................................................ 10 3.2 Descriptions of the Ecological Systems ............................................................................................................ 10 3.2.1 Spruce Forest: Central and Southern Appalachian Spruce-Fir Forest .......................................................... 10 3.2.2 Northern Hardwood Forest : Appalachian (Hemlock)- Northern Hardwood Forest ..................................... 11 3.2.3 Cove Forest: Southern and Central -

Assessment – Western Hemisphere Geography PP Eastern Hemisphere Geography Pre/Post Assessment
Assessment – Western Hemisphere Geography PP Eastern Hemisphere Geography Pre/Post Assessment GRADE 7 EASTERN HEMISPHERE GEOGRAPHY Pacing Calendar 2017-2018 Social Studies Process Skills for Organizing and Analyzing Information Resources, Websites and Activities Process Standards: Graphic Organizers Game Site: I Like to Learn 1. Identify, analyze, and interpret primary Interactive Graphic Organizers and secondary sources and artifacts. Graphic Organizers Print Outs Outline Maps 2. Distinguish between fact and opinion in examining documentary sources Organizing and Analyzing Information Nystrom Desk Atlas including the district website copy 3. Recognize and explain how different GIST Literary Selections/Books correlated to curriculum points of view have been influenced by History Frames (district purchase) History Frame Form social, political, economic, historic and History Frame Pyramid USA Test Prep Mind Maps Websites: 4. Construct timelines of key events, Mind Map Examples for Geography periods, and historically significant Mind Map Forms Mr Roughton Lesson Plans and Resources (New) individuals. One Sentence Summary TED Ed Lessons (New) 5. Explain the relationships between One Word Summary Geography from Tolland Middle School geography and the historical Power Thinking Chart for Geography developments by using maps, graphs, Prediction with Evidence Reynolds Geography Tolland Middle School charts, visual images, and computer- Problem-Solution Chart Think Pair Share Quizlet based technologies. Three Minute Pause Engaging Students with Primary Source Documents 6. Develop discussion, debate, and Three Minute Pause Chart persuasive writing and speaking skills, Analyzing an Historic Event District Geography Power Point Website focusing on enduring issues and APPARTS Geography Power Points demonstrating how divergent viewpoints Five Themes Analysis for Geography have been and continue to be addressed Five Themes Note-taking for Geography Geography Lesson Plans H Diagram Mr. -

5Th Grade Life Science: Ecosystems Unit
5th Grade Life Science: Ecosystems Unit Developed for Chapel Hill Carrboro City Schools Northside Elementary School Outdoor Wonder & Learning (OWL) Initiative Unless otherwise noted, activities written by: Lauren Greene, Sarah Yelton, Dana Haine, & Toni Stadelman Center for Public Engagement with Science UNC Institute for the Environment In collaboration with 5th grade teachers at Northside Elementary School: Michelle Gay, Daila Patrick, & Elizabeth Symons ACKNOWLEDGEMENTS Many thanks to Dan Schnitzer, Coretta Sharpless, Kirtisha Jones and the many wonderful teachers and support staff at Northside Elementary for their participation in and support of the Northside OWL Initiative. Thanks also to Shelby Brown for her invaluable assistance compiling, editing, and proofreading the curriculum. Instructional materials and supplies to promote STEM-based outdoor learning were instrumental to the successful implementation of this curriculum. The purchase of these materials was made possible with funding provided by the Duke Energy Foundation to Chapel Hill-Carrboro City Schools. Curriculum developed June 2018 – July 2019 For more information, contact: Sarah Yelton, Environmental Education & Citizen Science Program Manager UNC Institute for the Environment Center for Public Engagement with Science [email protected] 5th Grade Ecosystems Unit Northside Outdoor Wonder & Learning Initiative Overarching Unit Question How and why do organisms (including humans) interact with their environment, and what are the effects of these interactions? Essential Questions Arc 1: How can I describe and compare different ecosystems? Arc 2: How is energy transferred through an ecosystem? How can I explain the interconnected relationships between organisms and their environments? Transfer Goals o Use scientific thinking to understand the relationships and complexities of the world around them. -

Ecosystem Ecology Living Organisms and Their Non Living (Abiotic) Environment Are Interrelated and Interact with Each Other
1 | P a g e Ecosystem Ecology Living organisms and their non living (abiotic) environment are interrelated and interact with each other. The term ‘ecosystem’ was proposed by A. G. Tansley and it was defined as the system resulting from the integration of all living and non living factors of the environment. Some parallel terms such as biocoenosis, microcosm, geobiocoenosis, holecoen, biosystem, bioenert body and ecosom were used for each ecological systems. However, the term ‘ecosystem’ is most preferred, where ‘eco’ stands for the environment, and ‘system’ stands for an interacting, interdependent complex. Any unit that includes all the organisms, i.e. communities, in a given area, interacts with the physical environment so that a flow of energy leads to a clearly defined trophic structure, biotic diversity and material cycle within the system, is known as an ecological system or ecosystem. It is usually an open system with regular but variable influx and loss of materials and energy. It is a basic functional unit with unlimited boundaries. Types of ecosystems The ecosystems can be broadly divided into the following two types; 1. Natural ecosystems 2. Artificial (man cultured ecosystems Natural ecosystems The ecosystems which are self-operating under natural conditions with any interference by man, are known as natural ecosystems. These ecosystems may be classified as follows: 1. Terrestrial ecosystems, e.g. Forests, grasslands and deserts 2. Aquatic ecosystems, which may be further classified as i) Freshwater: ecosystem, which in turn is divided into lentic (running water like streams, springs, rivers, etc.) and lentic (standing water like lakes, bonds, pools, swamps, ditches, etc.) ii) Marine ecosystem, e.g. -

Distribution Mapping of World Grassland Types A
Journal of Biogeography (J. Biogeogr.) (2014) SYNTHESIS Distribution mapping of world grassland types A. P. Dixon1*, D. Faber-Langendoen2, C. Josse2, J. Morrison1 and C. J. Loucks1 1World Wildlife Fund – United States, 1250 ABSTRACT 24th Street NW, Washington, DC 20037, Aim National and international policy frameworks, such as the European USA, 2NatureServe, 4600 N. Fairfax Drive, Union’s Renewable Energy Directive, increasingly seek to conserve and refer- 7th Floor, Arlington, VA 22203, USA ence ‘highly biodiverse grasslands’. However, to date there is no systematic glo- bal characterization and distribution map for grassland types. To address this gap, we first propose a systematic definition of grassland. We then integrate International Vegetation Classification (IVC) grassland types with the map of Terrestrial Ecoregions of the World (TEOW). Location Global. Methods We developed a broad definition of grassland as a distinct biotic and ecological unit, noting its similarity to savanna and distinguishing it from woodland and wetland. A grassland is defined as a non-wetland type with at least 10% vegetation cover, dominated or co-dominated by graminoid and forb growth forms, and where the trees form a single-layer canopy with either less than 10% cover and 5 m height (temperate) or less than 40% cover and 8 m height (tropical). We used the IVC division level to classify grasslands into major regional types. We developed an ecologically meaningful spatial cata- logue of IVC grassland types by listing IVC grassland formations and divisions where grassland currently occupies, or historically occupied, at least 10% of an ecoregion in the TEOW framework. Results We created a global biogeographical characterization of the Earth’s grassland types, describing approximately 75% of IVC grassland divisions with ecoregions. -

Regions and People of the Eastern Hemisphere
6th Grade Social Studies Curriculum Map Regions and People of the Eastern Hemisphere FIRST SEMESTER Unit Strands and Content Statements Student Expectations Resources/Assessments Key Concepts 1 Geography Geography (3) Lesson Plan-Mapping Perceptions Basic terms www.asiasociety.org (3) Globes and other geographic tools (3) Use appropriate maps, globes and Basic Geography (3) National Atlas Map Maker tool can be used to gather, process and geographic tools to gather, process and Continents and Map Skills report information about people, http://www.nationalatlas.gov/mapmaker report information about people, places (4 weeks) places and environments. Longitude and and environments. Identify the seven continents and 5 Cartographers decide which oceans Latitude (revisit Big Idea: information to include and how it is (3) Understand that maps are created for in all units) specific purposes and represent the Types of maps and their purposes displayed. (physical, political, population density, Essential Questions: context in which they were created. climate, topographic, resource, Hemispheres How do we know historical) about the world (4) Latitude and Longitude can be (4) Use latitude and longitude coordinates Oceans (4) National Atlas (articles of latitude and today? used to identify absolute location to identify absolute location longitude) Types of Maps http://nationalatlas.gov/articles/mapping/a_l Political How does where atlong.html (5) Regions can be determined, Physical you live influence (5) Use various criteria to describe, classify (4) Crack the Code (solving mysteries using classified and compared using various and compare regions within the Eastern longitude and latitude) Topographic how you live? criteria (e.g. landform, climate, Climate Hemisphere. -

Effects on Ecosystems
10 Effects on Ecosystems J.M. MELILLO, T.V. CALLAGHAN, F.I. WOODWARD, E. SALATI, S.K. SINHA Contributors: /. Aber; V. Alexander; J. Anderson; A. Auclair; F. Bazzaz; A. Breymeyer; A. Clarke; C. Field; J.P. Grime; R. Gijford; J. Goudrian; R. Harris; I. Heaney; P. Holligan; P. Jarvis; L. Joyce; P. Levelle; S. Linder; A. Linkins; S. Long; A. Lugo, J. McCarthy, J. Morison; H. Nour; W. Oechel; M. Phillip; M. Ryan; D. Schimel; W. Schlesinger; G. Shaver; B. Strain; R. Waring; M. Williamson. CONTENTS Executive Summary 287 10.2.2.2.3 Decomposition 10.2.2.2.4 Models of ecosystem response to 10.0 Introduction 289 climate change 10.2.2.3 Large-scale migration of biota 10.1 Focus 289 10.2.2.3.1 Vegetation-climate relationships 10.2.2.3.2 Palaeo-ecological evidence 10.2 Effects of Increased Atmospheric CO2 and 10.2.2.4 Summary Climate Change on Terrestrial Ecosystems 289 10.2.1 Plant and Ecosystem Responses to 10.3 The Effects of Terrestrial Ecosystem Changes Elevated C02 289 on the Climate System 10.2.1.1 Plant responses 289 10.3.1 Carbon Cycling in Terrestrial Ecosystems 10.2.1.1.1 Carbon budget 289 10 3.1 1 Deforestation in the Tropics 10.2.1.1.2 Interactions between carbon dioxide and 10.3.1 2 Forest regrowth in the mid-latitudes of temperature 290 the Northern Hemisphere 10.2.1.1.3 Carbon dioxide and environmental 10 3.1.3 Eutrophication and toxification in the stress 290 mid-latitudes of the Northern Hemisphere 10.2.1.1.4 Phenology and senescence 291 10.3.2 Reforestation as a Means of Managing 10.2.1.2 Community and ecosystem responses to Atmospheric