Why Is DNA Double Stranded? the Discovery of DNA Excision Repair Mechanisms
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
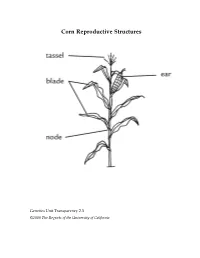
Corn Reproductive Structures
Corn Reproductive Structures Genetics Unit Transparency 2.3 ©2008 The Regents of the University of California Creating a Punnett Square Genetics Unit Transparency 2.4 ©2008 The Regents of the University of California Case Study Summary Sheet Case Study Type of Benefits Risks and concerns Remaining questions Genetic Modification Genetics Unit Transparency 4.1 ©2008 The Regents of the University of California Class Data for Rice Breeding Simulation aromatic, non-aromatic, aromatic, non-aromatic ,flood-tolear nt flood-tolerant flood-intolerant flood-intolerant AAFF AAFf AaFF AaFf aaFF aaFf AAff Aaff aaff Group 1 2 3 4 5 6 7 8 Genetics Unit Transparency 5.1 ©2008 The Regents of the University of California Three Types of Cells Genetics Unit Transparency 6.1 ©2008 The Regents of the University of California DNA base percentages in a variety of organisms Source of A T G C DNA Human 31.0% 31.5% 19.1% 18.4% Mouse 29.1% 29.0% 21.1% 21.1% Frog 26.3% 26.4% 23.5% 23.8% Fruit fly 27.3% 27.6% 22.5% 22.5% Corn 25.6% 25.3% 24.5% 24.6% E. coli 24.6% 24.3% 25.5% 25.6% Genetics Unit Transparency 7.1 ©2008 The Regents of the University of California Matthew Meselson and Franklin Stahl’s Experiment to Investigate DNA Replication First: Scientists James Watson and Francis Crick propose a method of semi-conservative replication in their paper on the structure of DNA. However, they have no data to support their hypothesis. Next: Matthew Meselson and Franklin Stahl use the procedure that follows to investigate DNA created by the process of DNA Replication. -

From the President's Desk
JAN/FEB 2006 From the President’s desk: 2006, the 75th anniversary of the Genetics Society of America, will be marked by a number of initiatives to reinvigorate the Society’s mission of promoting research and education in genetics. A highlight was the recently held GSA sponsored conference, “Genetic Analysis: From Model Organisms to Human Biology” in San Diego from January 5-7. This conference emphasized the importance of model organism research by illustrating the crucial contributions to human biology resulting from discoveries in these organisms. The National Institutes of Health (NIH) supported this conference both financially and by participation of key NIH administrators, including Jeremy M. Berg, director of the National Institute of General Medical Sciences. In addition to the superb science talks by international leaders the MOHB conference showcased other important and new GSA initiatives including education, public policy advocacy, graduate student support and recognition of outstanding model organism geneticists. Robin Wright, Education Committee chair, led a round table discussion on undergraduate education and the Joint Steering Committee for Public Policy and the Congressional Liaison Committee sponsored a session on science advocacy and public policy. There was a mentor lunch to support graduate students and postdocs in the next steps of their careers, and the three GSA medals were presented during the banquet, with Victor Ambros receiving the GSA Medal, Fred Sherman the Beadle Award, and Masatoshi Nei the Morgan Award. (For research highlights at the meeting, see pages 6 and 7 of this issue.) The 75th anniversary will also usher in changes to our society’s journal, GENETICS. -

Genetics Society News
July 2008 . ISSUE 59 GENETICS SOCIETY NEWS www.genetics.org.uk IN THIS ISSUE Genetics Society News is edited by Steve Russell. Items for future issues should be sent to Steve Russell, preferably by email to • Genetics Society Epigenetics Meeting [email protected], or hard copy to Department of Genetics, • Genetics Society Sponsored Meetings University of Cambridge, Downing Street, Cambridge CB2 3EH. The Newsletter is published twice a year, with copy dates of 1st June and • Travel, Fieldwork and Studentship Reports 26th November. • John Evans: an Appreciation Cocoons of the parasitoid wasp Cotesia vestalis on cabbage leaf in Taiwan. From the • Twelve Galton Lectures fieldwork report by Jetske G. de Boer on page 36. • My Favourite Paper A WORD FROM THE EDITOR A word from the editor ow soon until the $1000 based on the results of tests we genome is actually with barely understand! Here in the Hus and individual UK there is currently a sequencing is widespread? The moratorium, adhered to by publication of increasing most insurers, on the use of numbers of individual human genetic testing information for genome sequences suggests assessing life insurance that we should start to consider applications. It is important some of the implications that this remains in place and associated with the availability its effectiveness is reviewed of personal genetic well before the current information. In this issue we moratorium expires in 2011. present two articles reflecting The Human Genetics on his issue: a report from a Commission Genetics Society sponsored (http://www.hgc.gov.uk) meeting recently held in monitor issues relating to Cambridge organised by The genetic discrimination in the Triple Helix, an international UK and are a point of contact undergraduate organisation, as for those with any concerns in the Millennium Technology Prize. -

Issue 84 of the Genetics Society Newsletter
JANUARY 2021 | ISSUE 84 GENETICS SOCIETY NEWS In this issue The Genetics Society News is edited by • Non-canonical Careers: Thinking Outside the Box of Academia and Industry Margherita Colucci and items for future • Celebrating the 35th anniversary of DNA fingerprinting issues can be sent to the editor by email • Genetics Society Summer Studentship Workshop 2020 to [email protected]. • 2020 Heredity best student-led paper prize winners The Newsletter is published twice a year, • Industrious Science: interview with Dr Paul Lavin with copy dates of July and January. Celebrating students’ achievements: 2020 Genetics Society Summer Studentship Workshop, 2020 Heredity best student-led paper prize. Page 30 A WORD FROM THE EDITOR A word from the editor Welcome to Issue 84 elcome to the latest issue of the Thinking Outside the Box of WGenetics Society Newsletter! Academia and Industry”. This little This issue is packed with great news vade mecum for careers in genetics of achievements and good science. The collects inspiring interviews led first Genetics Society virtual workshop by our very own Postgraduate for the 2020 Summer studentship saw Representative, Emily Baker. In exceptional contributions from the Emily’s words, these experiences attending students. You can read more “demonstrate how a PhD in genetics about participants’ experiences in the can be a platform for a career in just interviews with the talk’s winners in about anything. Pursuing a career the Feature section. in academia, industry, publishing or science communication could be for Many more prizes were awarded: you, but so could many others. Why Heredity journal announced the not take a career path less travelled 2020 Heredity best student-led paper by, it might make all the difference?” winners, and James Burgon’s Heredity podcast dedicated an episode to the Enjoy! first prize winner, with insights from Best wishes, Heredity Editor-in-Chief, Barbara Margherita Colucci Mable. -

JESSICA L. MARK WELCH Assistant Research Scientist Josephine Bay
JESSICA L. MARK WELCH Assistant Research Scientist Josephine Bay Paul Center, Marine Biological Laboratory 7 MBL Street, Woods Hole, MA 02543 Telephone (508) 289-7180, e-mail [email protected] EDUCATION: B.A. magna cum laude, Harvard and Radcliffe Colleges, Biology, 1989 Graduate work, Freie Universität Berlin, Germany, Chemistry and Biology, 1990-1992 Advisor: Walter Sudhaus Ph.D., Harvard University Department of Molecular and Cellular Biology, Biology, 2001 Advisor: Matthew Meselson. Dissertation topic: Cytological Evidence for the Absence of Meiosis in Bdelloid Rotifers. Postdoctoral training, Marine Biological Laboratory, Woods Hole, Molecular Biology 2001-2006 part-time 2006-2009 full-time Advisors: Matthew Meselson and Gary Borisy PROFESSIONAL APPOINTMENTS: 1989 – 1990 Research Assistant, Environmental Law Institute, Washington, DC 2009 – present Assistant Research Scientist, Josephine Bay Paul Center for Comparative Molecular Biology and Evolution, Marine Biological Laboratory, Woods Hole, MA. PUBLICATIONS: Valm, Alex M., Jessica L. Mark Welch, and Gary G. Borisy (2012). CLASI-FISH: Principles of Combinatorial Labeling and Spectral Imaging. Systematic and Applied Microbiology 2012 Apr 21 (Epub ahead of print). PMCID: PMC3407316. Sukenik, Assaf, Ruth N. Kaplan-Levy, Jessica Mark Welch, and Anton Post (2011). Massive multiplication of genome and ribosomes in dormant cells (akinetes) of Aphanizomenon ovalisporum (Cyanobacteria). The ISME Journal 2011.128 (epub ahead of print, October 6, 2011). Valm, Alex M., Jessica L. Mark Welch, Christopher W. Rieken, Yuko Hasegawa, Mitchell L. Sogin, Rudolf Oldenbourg, Floyd E. Dewhirst, and Gary G. Borisy (2011). Systems-level analysis of microbial community organization through combinatorial labeling and spectral imaging. Proceedings of the National Academy of Sciences (USA) 108: 4152-4157. -

Die Molekularbiologie in Deutschland Von 1945 Bis 1975
Die Molekularbiologie in Deutschland von 1945 bis 1975 Ein internationaler Vergleich Inauguraldissertation zur Erlangung des Doktorgrades der Mathematisch-Naturwissenschaftlichen Fakultät der Universität zu Köln vorgelegt von Simone Wenkel aus Villingen-Schwenningen 2013 Die vorliegende Arbeit wurde am Institut für Genetik der Universität zu Köln in der Arbeitsgruppe für Geschichte der biologischen und chemischen Wissenschaften (Prof. Dr. rer. nat. Ute Deichmann) angefertigt. Berichterstatter: Prof. Dr. rer. nat. Ute Deichmann Prof. Dr. rer. nat. Thomas Wiehe Prüfungsvorsitz: Prof. Dr. rer. nat. Siegfried Roth Tag der mündlichen Prüfung: 24. Januar 2014 2 Zusammenfassung Die Molekularisierung der Biologie seit dem zweiten Drittel des 20. Jahrhunderts hatte immense Auswirkungen auf die Forschung und führte zu weitreichenden Anwendungen. Sie vereinte in einer Synthese viele biologische, biochemische und medizinische Disziplinen unter zentralen biologischen Fragestellungen. Durch die Entwicklung neuer Methoden und die Etablierung neuer Modellorganismen gelang es innerhalb weniger Jahrzehnte, die klassische Genetik, Mikrobiologie, Makromolekulare Chemie und Stoffwechselbiochemie miteinander in Verbindung zu bringen. In Deutschland war die Forschung nach 1945 viele Jahre lang geprägt von den Nachwirkungen der NS-Zeit und des Zweiten Weltkriegs, dem Wiederaufbau und der Neugründung von Instituten sowie großen Anstrengungen einzelner Wissenschaftler bei der Etablierung neuer Gebiete, wie dem der Molekularbiologie. Das Ziel dieser Arbeit ist es, erstmals ein umfassendes Bild der frühen Geschichte der Molkularbiologie in Deutschland zu erstellen und dieses im internationalen Vergleich zu betrachten. Zuerst wird die Entwicklung der Genetik und Molekularbiologie an deutschen Hochschulen und Forschungseinrichtungen im Hinblick auf die Institutionalisierung und Förderung analysiert. Neben der allgemeinen Entwicklung wird hier der Einfluss einzelner Personen, vor allem der des Physikers und Molekularbiologen Max Delbrück, herausgearbeitet. -

Matthew Meselson and Federal Policy on Chemical and Biological Warfare
142 The People's Science Advisora-Can Outsiders Be Ef(ectivef . D .__ 30 1969 p 14· January 4, 1970, p. 29; February 2, 53. New York T1me1, ecem..... • ' • ' CHAPTER 11. 1970,p.27. 35 54 New York Timei,June 18, 1970, p. • 0 6 9 55: Time, April 12, 1971, P· 45;New YorkNT::J~~~~! i?e,i!~s ·in· Pepper Fields," 56. Robert Gillette, "DDT: lts Days ue u er ' Seien« 176 (1972): 1313. 0 72-1548 etc. (6 suits). December 18, 197~. " Matthew Meselson and the 51. EDF et. al v. EPA etc., D.C. Clorar.. p. Result of lnsect Resistance, New Chemu:als, 58. "Decreasing Use of Organoch mes as 9 Chemical and Engineering N;,wsk, AT~~ No• ~:~~·{1\911, Section IV, P· 13. 59. Quoted in the New • or rme-, • United States Policy on 60. Jbid. 61. Jbid. ered •• 1314 62. Gillette, "DDT: lts Days Are Numb ••AiXrin and Dieldrin," Environment, October 63. See, for example, tbarles F. Wurstel, Chemical and 1971, p. 33. • . .cals re ·stered ror use in the United States. 64. In 1969 there were 900 pestic:'dal c~ on';.e1ticide111nd Their Relationship to See, c.g., thc Report of the SeaetflTY 'Com n Biological Warfare Envi1onmento1Health, p. 46. Mattbew Meselson is a sligbt, soft-spoken professor of biochemistry at Harvard wbo often seems to be occupying the calm at the center of a burricane of activity. The scene wbicb greeted one of the authors on an aftemoon visit to bis laboratory during the spring of 1973 was typical: Meselson's graduate students bad congregated for wine, cheese, discussion, and laughter in a room next to bis office. -

100 Years of Genetics
Heredity (2019) 123:1–3 https://doi.org/10.1038/s41437-019-0230-2 EDITORIAL 100 years of genetics Alison Woollard1 Received: 27 April 2019 / Accepted: 28 April 2019 © The Genetics Society 2019 The UK Genetics Society was founded on 25 June 1919 and “biometricians”; the Genetical Society was very much a this special issue of Heredity, a journal owned by the society of Mendelians. Remarkably, 16 of the original 87 Society, celebrates a century of genetics from the perspec- members were women—virtually unknown in scientific tives of nine past (and present) presidents. societies at the time. Saunders was a vice president from its The founding of the Genetical Society (as it was then beginning and its 4th president from 1936–1938. Perhaps known) is often attributed to William Bateson, although it the new, and somewhat radical, ideas of “genetics” pre- was actually the brain child of Edith Saunders. The enthu- sented a rare opportunity for women to engage in research siasm of Saunders to set up a genetics association is cited in because the field lacked recognition in universities, and was the anonymous 1916 report “Botany at the British Asso- therefore less attractive to men. 1234567890();,: 1234567890();,: ciation”, Nature, 98, 2456, p. 238. Furthermore, the actual Bateson and Saunders (along with Punnett) were also founding of the Society in 1919 “largely through the energy influential in the field of linkage analysis (“partial coupling” as of Miss E.R Saunders” is reported (anonymously) in they referred to it at the time), having made several observa- “Notes”, Nature, 103, 2596, p. -

Issue 82 of the Genetics Society Newsletter
JANUARY 2020 | ISSUE 82 GENETICS SOCIETY NEWS In this issue The Genetics Society News is edited by Margherita Colucci and items for future • Medal and Prize Lecture Announcements issues can be sent to the editor by email • “A Century of Genetics” conference to [email protected]. • Celebrating the centenary of Fisher 1918 The Newsletter is published twice a year, • Research and travel grant reports with copy dates of July and January. Speakers’ dinner at the “A Century of Genetics” conference, November 2019, Edinburgh. (Photo by Douglas Vernimmen) A WORD FROM THE EDITOR A word from the editor Welcome to Issue 82 elcome to the latest issue of reports in the Sectional Interest Wthe GenSoc Newsletter and Groups: Reports section. my first steps (pages?) as new editor. And why not (re)discovering another I am eager to start this journey with great milestone such as the publishing you through the latest Genetics of Fisher’s 1918 paper, “The correlation Society achievements and genetics between relatives on the supposition news! I would like to thank all of Mendelian inheritance”, recently GenSoc committee for giving me this reaching its centenary recurrence? opportunity. I am sure you will greatly enjoy the In this issue, I will bring you back to report in the Features section. the inspiring and lively atmosphere Enjoy! of the GenSoc meeting ‘A Century of Genetics’ in Edinburgh (November Best wishes, 2019) - a really big thanks to all of those Margherita Colucci who kindly contributed. Many Sectional Interest groups have been very active: you will find their In this issue, I will bring you back to the inspiring and lively atmosphere of the GenSoc meeting “A Century of Genetics” in Edinburgh (November 2019) - a really big thanks to all of those who kindly contributed. -

Allele Sharing and Evidence for Sexuality in a Mitochondrial Clade of Bdelloid Rotifers
HIGHLIGHTED ARTICLE GENETICS | INVESTIGATION Allele Sharing and Evidence for Sexuality in a Mitochondrial Clade of Bdelloid Rotifers Ana Signorovitch,* Jae Hur,*,† Eugene Gladyshev,* and Matthew Meselson*,‡,1 *Department of Molecular and Cellular Biology, Harvard University, Cambridge, Massachusetts 02138, †Department of Biology, Harvey Mudd College, Claremont, California 91711, and ‡Josephine Bay Paul Center, Marine Biological Laboratory, Woods Hole, Massachusetts 02543 ORCID IDs: 0000-0003-1446-2305 (J.H.); 0000-0002-4074-4966 (E.G.); 0000-0002-9827-2682 (M.M.) ABSTRACT Rotifers of Class Bdelloidea are common freshwater invertebrates of ancient origin whose apparent asexuality has posed a challenge to the view that sexual reproduction is essential for long-term evolutionary success in eukaryotes and to hypotheses for the advantage of sex. The possibility nevertheless exists that bdelloids reproduce sexually under unknown or inadequately investigated conditions. Although certain methods of population genetics offer definitive means for detecting infrequent or atypical sex, they have not previously been applied to bdelloid rotifers. We conducted such a test with bdelloids belonging to a mitochondrial clade of Macrotrachela quadricornifera. This revealed a striking pattern of allele sharing consistent with sexual reproduction and with meiosis of an atypical sort, in which segregation occurs without requiring homologous chromosome pairs. KEYWORDS clonal erosion; cyclic parthenogenesis; meiosis; monogonont; Oenothera DELLOID rotifers are minute freshwater invertebrates com- families and hundreds of morphospecies, their ancient origin is Bmonly found in lakes, streams, and ponds and in ephem- indicated by the considerable synonymous sequence divergence erally aquatic environments such as rock pools and the water between families and by the finding of bdelloid remains in 35- to films on moss and lichens where they are able to survive because 40-million year old amber (Mark Welch et al. -

Perspectives
Copyright Ó 2007 by the Genetics Society of America Perspectives Anecdotal, Historical and Critical Commentaries on Genetics Edited by James F. Crow and William F. Dove R. A. Fisher’s 1943 Unravelling of the Rhesus Blood-Group System A. W. F. Edwards1 Gonville and Caius College, Cambridge CB2 1TA, United Kingdom VEN if R. A. Fisher’s elucidation of the human blood- binatorial skills and his relations with A. S. Wiener, and E group system Rhesus in terms of the three linked to add a personal coda. It is best to take first the diagram loci C, D, and E had not proved to be substantially (Figure 1) from Fisher (1947) because it explains the correct, it would still have been an outstanding example relationship between the original Rhesus notation and of the power of analytical thought to unravel a complex that proposed by Fisher. As Fisher writes, ‘‘We may repre- array of genetical data. In fact, as a recent review relates sent the eight heritable antigen complexes geometrically (Avent et al. 2006), D is one gene (carrying the D anti- as the corners of a cube, while the six elementary antigens gen) and C and E are different splicing forms of another are represented by the faces; each allelomorphic pair of (CE carrying the C or c antigens and the E or e antigens). antigens is then a pair of opposite faces, and the three Fisher’s solution is recognizable beneath the modern faces meeting in any point specify the antigens in each molecular detail. complex.’’ (Ry) and (CdE) are in parentheses because The story of the unravelling of the Rhesus puzzle is the anti-d antibody had not yet been discovered (nor has told in chapter 13 of Joan Fisher Box’s R. -

Genetics Society of America
Genetics Society of America 1992 Records, Proceedings and Reports Published as supplementary material in GENETICS, Volume 131 Prepared by The Secretary Shirleen Roeder Department of Biology Yale University New Haven, Connecticut s2 BOARDS, COMMITTEES AND REPRESENTATIVES FOR 1992 BOARD OF DIRECTORS John C. Lucchesi, President Karen Artzt A. Dale Kaiser, Vice-president Welcome Bender Shirleen Roeder, Secretary Nina V. Fedoroff Carol S. Newlon, Treasurer Barry S. Ganetzky Leland H. Hartwell, Past President Christine Guthrie John W. Drake, Editor H. Robert Horvitz GENETICSEDITORIAL BOARD John W. Drake, Editor Maureen R. Hanson Sally Lyman Allen Robert K. Herman Karen Artzt Alan G. Hinnebusch Douglas E. Berg Richard R. Hudson David Botstein Nancy A. Jenkins John E. Boynton Mark Johnston Anthony H. D. Brown Elizabeth Jones Benjamin Burr Cathy C. Laurie Marian Carlson Wen-Hsiung Li Deborah Charlesworth Trudy F. C. Mackay Peter Cherbas Patricia J. Pukkila Arthur Chovnick Trudi Schiipbach Andrew G. Clark William F. Sheridan Thomas W. Cline Michael J. Simmons Rowland H. Davis Montgomery Slatkin Robin E. Denell Gerald R. Smith Norman R. Drinkwater Steven D. Tanksley Walter F. Eanes Elizabeth Thompson Victoria G. Finnerty Bruce S. Weir Margaret T. Fuller Fred Winston Roger E. Ganschow ADMINISTRATIVE OFFICE Elaine Strass, Executive Director Barbara Abbott, Publicat~nsAssistant/ASHG Membership Coordinator Sharon Adler, Accounting Assistant Judy Ashton, Oflce Coordinator Candis Galkin, Accounting Assistant Gloria Garber, Membership and Meetings Assistant Karen Gould, Comptroller/Manager of Administration Margot Kiley, GSA Membership Coordinator Krista Koziol, Publications Manager Anne Marie Langevin, GSA Meetings Manager Marsha Ryan, ASHG Meetings Manager Jane Salomon, ASHG Special Projects Manager COMMITTEES Leland H.